
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
Психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Сочинение: Исследование расщепления крахмала под действием a-амилазы слюны
Сочинение: Исследование расщепления крахмала под действием a-амилазы слюны
Общеобразовательная средняя школа №5
Исследовательская работа
Тема: «Исследование расщепления крахмала под действием a-амилазы слюны»
Авторы: Учитель химии Новиков В.В.
Ученицы 11а класса
Рогова Екатерина, Белова Юля
Содержание
2. Цель работы.. 3
3. Литературный обзор. 4
3.1. Крахмал. Строение крахмала. 4
3.2. Общие понятия о ферментах. 9
3.3. Характеристика амилаз. 12
3.4. a-Амилазы.. 13
3.5. Гидролиз крахмала a-амилазами. 14
4. Ход работы.. 16
5. Литература. 18
6. Благодарности. 19
1. Актуальность работы
Амилазы широко используются в пищевой промышленности. Так амилазы используются в хлебопечении и технологиях брожения. Также a-амилаза играет значительную роль в расщеплении крахмала в организме человека. Поэтому понимание действия амилазы важно для оптимизации промышленного производства и изучения обмена веществ в организме человека.
2. Цель работы
Исследовать активность фермента a-амилазы слюны при различных температурах и нахождение оптимальной температуры действия фермента в нейтральной среде.
3. Литературный обзор
3.1. Крахмал. Строение крахмала
Крахмал – один из наиболее распространенных запасных полисахаридов растений. Он интенсивно накапливается в результате фотосинтеза и откладывается в семенах, клубнях и других частях растений. Семена и клубни содержат 40-70% крахмала, другие части растений от 4 до 25%. При кислотном гидролизе крахмал распадается с образованием D - глюкозы, являющейся его структурным элементом, и небольшого количества глюкозо-6 - фосфата, так как все виды крахмала содержат немного (0,02 – 0,16%) фосфора. Установлено, что глюкоза в составе крахмала находится в виде α – D – глюкопиранозы.
Природный крахмал состоит из двух различных фракций, отличающихся по своему строению и свойствам. Примерно 20% крахмала составляет амилоза (от греч. амилон–крахмал). Остальное приходится на вторую фракцию, получившую название амилопектина (от греч. пектос – студнеобразный). Такая терминология отражает некоторые свойства этих двух видов крахмала. Амилопектин с трудом растворяется в горячей воде, причём раствор получается вязкий (крахмальный клейстер) и при охлаждении застывает в студневидную массу. Амилоза же хорошо растворима в тёплой воде и не образует клейстера. Пользуясь этим обстоятельством, амилозу отделяют от амилопектина, многократно извлекая её тёплой водой. С этой же целью используют способность амилозы осаждаться под действием бутилового спирта при насыщении последним горячего раствора, содержащего смесь амилозы и амилопектина. Применяют и хроматографические методы. Например, после пропускания диспергированного крахмала через колонку с фосфатом кальция и последующего промывания фосфатным буфером амилоза элюируется, а амилопектин остаётся на сорбенте.
Молекулярная масса амилозы и амилопектина различна: у не деградировавшиз в процессе выделения препаратов амилозы она составляет от 100000 до 400000, а у амилопектина превышает, как правило 20*106. Соответственно коэффициент полконденсации α-D-глюкопиранозы в молекулах амилозы оценивается в несколько сотен, а у амилопектина – в несколько десятков и даже сотен тысяч.
Различна и химическая структура амилозы и амилопектина. Молекулы первой, как правило, строго линейны. В них остатки a-D-глюкопиранозы связаны друг с другом исключительно a-1,4-глюкозидными связями, т.е. кислородные мостики возникают за счёт гликозидного гидроксила 1-го атома одной молекулы a-D-глюкопиранозы и спиртового гидроксила при 4-м атоме другой:
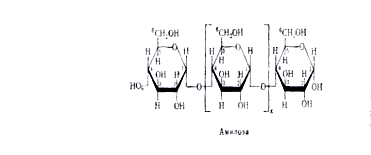
В соответствии с таким строением амилозу можно характеризовать как a-1,4-глюкан. Таким образом, амилоза представляет линейный полисахарид, молекулы которого имеют нитевидную структуру.
Современные данные о строении амилозы основаны на предположении, что остатки a-D-глюкопиранозы в её составе в силу некоторых обстоятельств имеет конформацию типа лодки. В этом случае структурная формула амилозы принимает следующий вид:

Лодкообразная конформация a-D - глюкопиранозных остатков в молекуле амилозы способствует спирализации полигликозидной цепи. При этом один виток спирали включает 6-7 остатков глюкозы. При длине каждого остатка глюкозы, равной 0,5 нм, возникает спираль диаметром 1 нм. Допускают, что молекулы амилозы, как и других линейных полисахаридов, могут на том или ином протяжении взаимодействовать друг с другом, образуя вторичные структуры бисприрального типа с взаимно закрученными полисахаридными цепями.
Амилопектин имеет сферические молекулы с радиусом вращения от 82 до 255 нм. Их сферическая форма обеспечивается тем, что молекула составлена из множества (несколько сотен) коротких полигликозидных цепочек, каждая из которых в среднем содержит 20 остатков a-D - глюкопиранозы. В пределах каждой короткой цепи глюкозные остатки соединены a-1,4 гликозидными связями. Друг с другом цепи соединяются посредством a-1,6 гликозидных связей. Строение этого разветлённого участка молекулы амилопектина таково:
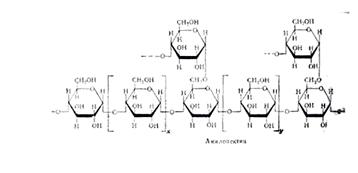
Общая структура молекулы амилопектина в соответствии с ранними данными показана на рис. На этом же рисунке приведена более современная модель амилопектина, выведенная на основании детального исследования продуктов ферментативного гидролиза и рентгенографического анализа этого полисахарида. Она получила название гроздевидной, так как по расположению в ней полигликозидных звеньев весьма напоминает гроздь винограда. Степень полимеризации остатков a-D-глюкозы в звеньях, обозначенных утолщёнными линиями, достигает 45, тонкими – 15. Протяжённость каждой псевдокристаллической области составляет 6 нм, аморфой – втрое меньшую величину
Считают, что в составе амилопектина a-D-глюкопираноза также находится в лодкообразной конформации. Вследствие этого отдельные участки полигликозидных цепочек, составляющих молекулу амилопектина, видимо спирализованы подобно амилозе.
Обе фракции крахмала дают окрашивание с йодом в растворе KI, однако амилоза окрашивается в чисто синий цвет, а амилопектин – в фиолетовый. Реакция крахмала с йодом не связана с химическим взаимодействием между ними, а состоит в образовании комплексов адсорбционного типа. Так как ведущую роль в возникновении этих комплексов играют спиральные участки молекул крахмала того или другого типа, то различие в тональности окраски вполне естественно. Предполагают, что молекулы йода втягиваются внутрь полигликозидной спирали, где и замыкаются соответствующие связи, дающие начало цветным комплексам.
При кратковременном нагревании порошкообразного крахмала его гигантские молекулы распадаются, образуя смесь более простых полисахаридов меньшей молекулярной массы – декстринов. Декстринизация крахмала при нагревании сопровождается повышением его растворимости в воде. Обработанный таким образом крахмал называют растворимым. Распад молекул крахмала до декстринов особенно интенсивно идёт при нагревании крахмального клейстера с 10% -ным раствором H2SO4. При дальнейшей обработке молекулярная масса декстринов прогрессивно падает и конечным продуктом распада является D-глюкоза. Крупномолекулярные декстрины окрашиваются йодом в красный цвет, низкомолекулярные – окраски с йодом не дают. Таким образом, гидролиз крахмала происходит ступенчато.
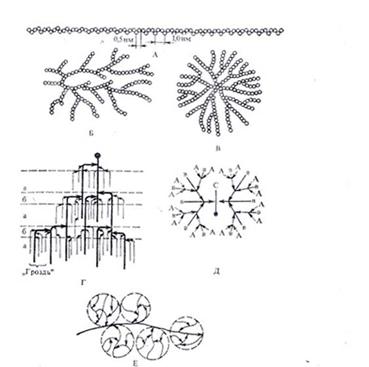
Рис.1 А – амилоза; Б – амилопектин; В – гликоген; каждый кружок обозначает остаток глюкозы; Г – современная гроздевидная модель молекулы амилопектина (а – компактная, псевдокристалическая область, б – менее компактная аморфная область; точкой обозначена редуцирующая группа); Д – модернизированная модель молекулы гликогена, демонстрирующая наличие спрятанных полигликозидных цепей; Е - строение частичкового гликогена; центральная линия – полипептидная цепь, к которой через остатки фосфата присоединены субъединицы гликогена.
3.2. Общие понятия о ферментах
Важнейшим свойством ряда белков является их каталитическая активность. Вещества белковой природы, способные каталитически ускорять химические реакции, называют ферментами (от лат. Fermentum – закваска) или энзимами (от греч. ен – внутри, зим – закваска). Как вытекает из происхождения названий этих веществ, первые сведения об их существовании были получены при изучении процессов брожения.
Роль ферментов в жизнедеятельности животных, растений и микроорганизмов колоссальна. Благодаря каталитической функции разнообразные ферменты обеспечивают быстрое протекание в организме или вне его огромного числа химических реакций. Складываясь в единый ансамбль саморегулируемых биохимических процессов, эти реакции преобразования веществ составляют материальную и энергетическую основу непрерывного самообновления белковых тел, т.е. самой сущности жизненных явлений. Поэтому ферменты «есть возбудители всех химических превращений». (И.П. Павлов)
В настоящее время в биологических объектах обнаружено несколько тысяч индивидуальных ферментов, а несколько сотен из них выделено и изучено. Подсчитано, что живая клетка может содержать до 1000 различных ферментов, каждый из которых ускоряет ту или иную химическую реакцию.
Биологические катализаторы (ферменты) по ряду признаков резко отличаются от неорганических катализаторов, хотя и те и другие лишь ускоряют достижение равновесия в химических процессах, которые протекают сами по себе, но сочень малыми скоростями. Как и катализаторы неорганической природы, биокатализаторы не вызывают каких либо химических реакций, а лишь ускоряют существующие. Первое различие состоит в том, что по сравнению с катализаторами неорганической природы ферменты «работают» в очень мягких условиях (низкая температура, нормальное давление, невысокие значения pH среды) и очень интенсивно. Так, например, гидролитический распад белка до аминокислот в присутствии неорганических катализаторов(крепких кислот и щелочей) осуществляется при температуре 100ºС и выше за несколько десятков часов. Этот же процесс при каталитическом участии специфических ферментов требует всего несколько десятков минут и идёт при температуре 30-40ºС. Для гидролиза крахмала, как указывал ещё Й. Берцелиус (1836), при нагревании в растворе кислоты нужно несколько часов, а при участии соответствующего фермента этот процесс идёт при комнатной температуре и занимает всего несколько минут. Ионы Fe каталитически ускоряют разложение H2O2 на Н2О и О2. Однако атомы того же Fe, но в составе фермента каталазы действуетв 10 млрд. раз энергичнее, и всего 1 мг Fe в ферменте способен заменить в этой реакции 10 т неорганического Fe. Таким образом, исключительно высокая каталитическая активность, проявляемая в условиях нормальной температуры и давления, отличает биокатализаторы от неорганических катализаторов.
Второе различие заключается в том, что ферменты обладают необыкновенно высокой специфичностью действия, чего не наблюдается у катализаторов неорганической природы. Каждый фермент каталитически ускоряет, как правило, одну-единственную химическую реакцию или в крайнем случае группу реакций одного типа.
Наконец, ряд различий между биокатализаторами и неорганическими катализаторами связан с белковой природой ферментов. Сюда относится термолабильность, зависимость активности от pH и наличие активаторов или ингибиторов и др.
Самая существенная разница между ферментами и обычными катализаторами вскрыта лишь в последние годы. Она состоит в том, что благодаря уникальной структуре каждого фермента процесс ферментативного катализа представляет перед нами как серия элементарных превращений вещества, строжайшим образом организованным в пространстве и во времени. Кооперативность и жёсткая запрограммированость действия – вот что отличает механизм биокатализа от действия катализаторов иной природы, хотя это не исключает некоторой степени вариабельности как структуры самого фермента, так и строения промежуточных продуктов в процессе ферментативного катализа.
В природе под каталитическим воздействием ферментов осуществляются процессы гидролиза, фосфоролиза, переноса различных групп (метильные радикалы, остатки фосфорной кислоты и т.д.) окисления и восстановления, расщепления и синтеза, изомеризации и т.п. Практически все химические преобразования в живом веществе протекают с помощью ферментов. Естественно поэтому, что каталитическая функция ферментов лежит в основе жизнедеятельности любого организма. При посредстве ферментов реализуется влияние как внутренних, генетических, так и внешних, природных факторов на развитие организма. Благодаря контакту ферментов с лекарственными веществами и антибиотиками достигается такое изменение ферментативных процессов, которые способствуют излечению от болезней, в то же времяизменение ферментативной активностипод влиянием микробных токсинов и иных ядов ведёт к гибели организма. Стимуляция роста животных и растений разнообразными препаратами, применяемые в сельском хозяйстве, в большинстве случаев основана на их воздействии, на процесс биосинтеза или активность тех или иных ферментов. Тончайшие различия строения ряда ферментов определяют видовые особенности организмов, а в нарушении биосинтеза некоторых из них заложена причина возникновения наследственных и других заболеваний. Всё это свидетельствует об огромном значении ферментов для биологии, сельского хозяйства и медицины.
Будучи выделены из организма, ферменты не утрачивают способность осуществлять каталитическую функцию. На этом основано их практическое применение в химической, пищевой, лёгкой и фармацевтической промышленности. Особое значение для химического производства имеет специфичность ферментов: ведь до 80% затрат в производстве многих химических веществ приходится на отделение примесей, возникающих в результате побочных реакций. Проведение синтеза при посредстве высокоспецифичного фермента, ускоряющего только ту реакцию, которая ведёт к образованию нужного продукта, упрощает технологический процесс. Кроме того, ферменты позволяют осуществлять ряд процессов, выполнение которых обычными методами органического синтеза остаётся пока нерешённой проблемой. Так обстоит дело, например, при получении лекарственных препаратов путём ферментативной трансформации стероидов.
3.3. Характеристика амилаз
Использование амилаз человеком было известно с незапамятных времён. Однако их изучение началось с открытия Кирхгофом в 1814 г. Вещества способного превращать крахмал в сахар. Препарат, полученный Кирхгофом из пшеничной муки, обладал способностью разжижать крахмальный клейстер и превращать его в сахарный сироп. Аналогичное явление автор наблюдал при смешивании крахмального клейстера с ячменным солодом. Уже в этих первых исследованиях Кирхгоф отметил губительное действие на это вещество серной кислоты. В то же время он подчеркнул, что сахарообразование является необходимым условием для сбраживания крахмалосодержащих материалов, и таким образом положил начало научному объяснению технологии брожения.
Биологически активное вещество в дальнейшем было выделено из солода и подверглось детальному изучению. Было обнаружено, что его действие на крахмал происходит через три стадии: разжижение, декстринизацию и осахаривание. Это привело к признанию существования в солоде двух различных компонентов, которые в дальнейшем были получены в отдельности и названы a-амилазой (декстринирующий компонент) и b-амилазой (осахаривающий компонент).
a-Амилазы и b-амилазы широко распространены в высших растениях. Наиболее важным источником амилаз являются хлебные злаки, зерно которых в проросшем состоянии (в виде солода) находит широкое применение в промышленном гидролизе крахмала. Солод из ячменя, ржи, пшеницы, овса, проса в настоящее время используются для осахаривания крахмала в спиртовом производстве.
3.4. a-Амилазы
Роль a-амилаз при гидролизе крахмала исключительно велика. Из трёх основных функций при действии на клейстеризованный крахмал (разжижение, декстринизация, осахаривание) разжижение и декстринизация зависят от a-амилаз. Они атакуют не только клейстеризованный, но и нативный крахмал, разрушая крахмальные зёрна.
a-Амилазы различного происхождения имеют много общих свойств: хорошо растворяются в воде или в сильно разбавленных растворах солей. Более концентрированные растворы солей (например 20-30% - ные растворы сульфата аммония) вызывают осаждение этих ферментов. a-Амилазы легко растворяются в разбавленных растворах этилового спирта, но осаждаются при его концентрации в среде свыше 60%. Белок a-амилаз обладает слабокислыми свойствами; изоэлектрическая точка ферментов колеблется в пределах рН 4,2 – 5,7. Молекулярная масса солодовой a-амилазы 60000, a - амилаз микроскопических грибов – 45000-50000. Многие из известных a-амилаз получены либо в высокоочищенном, либо в кристаллическом виде.
Ионы кальция оказывают стабилизирующее действие на a-амилазы. Это впервые было обнаружено Воллерштейном, затем потверждено Накамурой. В настоящее время это явление отмечено почти для всех амилаз. Однако теоретически этот вопрос применительно к промышленному гидролизу крахмала до сих пор не разработан.
3.5. Гидролиз крахмала a-амилазами
a-Амилазы действуют на a-1,4 –гликозидные связи, расщепляют амилозу внутри её цепи, в результате чего с большой скоростью образуются низкомолекулярные продукты гидролиза – нормальные a-декстрины. Их дальнейший гидролиз даёт мальтозу, мальтотриозу и глюкозу. Было найдено, что расщепление a-1,4-глюкозидных связей в амилозе носит случайный характер иподчиняется закону статистического распределения продуктов реакции. Расщепление более мелких фракций на последнем этапе амилоза носит уже не случайный характер – действие фермента направлено лишь на определённые a-1,4-гликозидные связи. В конечном счёте a-амилазы превращают амилозу в мальтозу и глюкозу, хотя и отмечены некоторые несущественные различия в динамике гидролиза этими ферментами указанного субстрата.
Бендецкий, Яровенко по изменению вязкости и восстанавливающей способности гидролизатов крахмала оценивали действие (множественность атаки) a-амилазы Bac. Subtilis на растворимый крахмал. Авторы наблюдали существенное различие вязкость –восстанавливающая способность при кислотном и ферментативном гидролизе крахмала. Это дало основание сделать заключение, что при кислотном гидролизе деградация крахмала происходит беспорядочно, а при действии a-амилазы осуществляется множественная атака на субстрат, приводящая к образованию олигомеров на первой стадии деградации.
Разрыв цепей амилопектина осуществляется между a-1,6 - глюкозидными связями. Отщепление декстринов, содержащих 15 и более гликозидных остатков, идёт с большей скоростью, в то время как конечное осахаривание существенно замедляется. Продукты гидролиза, содержащие a-1,4 - глюкозидные связи, являются нормальными a-декстринами и состоят из 6-13 глюкозидных остатков. Остаточные декстрины, содержащие большое количество a-1,6-связей, обазначаются как аномальные конечные декстрины. Установлено, что a-1,6-связи не только не расщепляются a-амилазой, но и являются стерическим препятствием для гидролиза a-1,4-связей, находящихся в непосредственной близости к a-1,6-связям. Наименьшим предельным декстрином в конце гидролиза амилопектина амилазой слюны был найден тетрасахарид, солодовой амилазы – паноза, т. е устойчивым к расщеплению в первом случае были две a-1,4-глюкозидные связи, во втором – лишь одна.
На рис. Представлена схема действия a-амилаз на амилозу и амилопектин по Бернфельду.
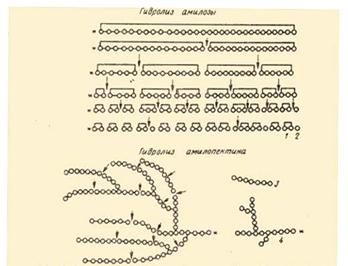
![]() Рис.2 Действие a-амилазы на амилозу и амилопектин по
Бернфельду.1 – мальтоза; 2 – глюкоза; 3 – нормальный a-декстрин; 4 – конечный a-декстрин;
*-редуцирующий конец амилозы или амилопектина; - действие a-амилазы.
Рис.2 Действие a-амилазы на амилозу и амилопектин по
Бернфельду.1 – мальтоза; 2 – глюкоза; 3 – нормальный a-декстрин; 4 – конечный a-декстрин;
*-редуцирующий конец амилозы или амилопектина; - действие a-амилазы.
4. Ход работы
Оборудование: электрическая плитка, химические стаканы, колбы, термостойкая колба, термометр, водяная баня, пипетка, мензурка.
Реактивы: крахмал, раствор йода.
Приготовление раствора крахмала:
В термостойкую колбу добавляем 200 мл воды. В неё же добавляем 2 г крахмала. Доводим смесь до кипения и кипятим несколько минут. Таким образом получаем 1% раствор крахмала.
Приготовление раствора слюны:
50 мл воды полощем во рту в течении 3-4 минут.
Проведение эксперимента:
В три пробирки добавляем по 25 мл раствора крахмала и каплю йода. Раствор крахмала окрашивается в интенсивно синюю окраску. Нагреваем электрическую плитку и измеряем температуру водяной бани. В течении 8-10 минут даём время, чтобы температура водяной бани и температура раствора в пробирках уравнялась. Снова измеряем температуру водяной бани. Добавляем в раствор крахмала 5 мл раствора слюны и включаем секундомер. Отмечаем время исчезновения синей окраски. Затем в следующие три пробирки добавляем раствор крахмала и каплю раствора йода, охлаждаем водяную баню и повторяем процедуру при более низкой температуре.
Результаты эксперимента приведены в таблице:
| toC | 22 | 30 | 34 | 37 | 40 | 45 | 58 | |
| t1 | 11 мин 17 с | 7 мин 5с | 3 мин 56 с | 3 мин 16 с |
4 мин 6 с |
5 мин 13 с |
6 мин 57 с |
|
| t2 | 11мин 22 с | 7 мин 13с |
4 мин 6 с |
3 мин 23 с |
4 мин 16 с |
5 мин 5 с |
6 мин 55 с | |
| t3 |
11 мин 23 с |
7 мин 9с | 4 мин |
3 мин 21 с |
4 мин 8 с |
5 мин 11 с |
7 мин 3 с |
|
| tср. | 11 мин 21с |
7 мин 9 с |
4 мин 1 с |
3 мин 20 с | 4 мин 10 с | 5 мин 10 с |
6 мин 58 с |
|
| toC | 22 | 30 | 34 | 37 | 40 | 45 | 58 | |
| 1/t мин-1 | 0.088 | 0.14 | 0.25 | 0.3 | 0.24 | 0. 195 | 0.143 | |
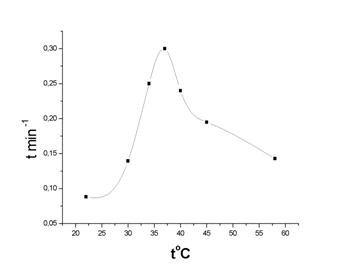
Зависимость скорости расщепления крахмала от температуры.
Можно видеть, что кривая имеет колокообразный характер с максимумом при температуре 37оС. Это подтверждает белковое строение a-амилазы, так как при использовании неорганических катализаторов скорость при повышении температуры постоянно возрастает. Также можно видеть, что в нейтральной среде оптимальная работа фермента происходит при 37оС, что совпадает с температурой тела человека.
5. Литература
1. Алейникова Т.Л., Рубцова Г.В. «Руководство к практическим занятиям по биологической химии». М.: Высшая школа 1988 г.
2. Жеребцов Н.А. «Амилолитические ферменты в пищевой промышленности». М.: Лёгкая и пищевая промышленность 1984 г.
3. А. Ленинджер «Биохимия». 1976 г.
4. Лещук Р.И., Хило З.В. «Малый практикум по биохимии». Томск 1984 г.
5. Производство и применение амилолитических ферментов в пищевой промышленности. М.: Пищевая промышленность 1968 г.
6. Филиппович Ю.Б. «Основы биохимии». М.: Высшая школа 1985 г.
6. Благодарности
Выражаю благодарность учителю физики Барановой Елене Львовне за помощь в работе и за предоставленное оборудование, ученику 9б класса Косареву Славе за оформление плакатов и ученице 11а класса Овчаровой Наталье за помощь в работе.