
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
Психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Реферат: Генетика и биометрия
Реферат: Генетика и биометрия
ФГОУ ВПО «Чувашская государственная
сельскохозяйственная академия»
Контрольная работа
по дисциплине
«Генетика и биометрия»
выполнила студентка
3 курса
заочного обучения
по специальности «Зоотехния»
Кудрявцева Екатерина Сергеевна
Шифр 08217
г. Чебоксары 2010г.
Содержание
1.Основные типы взаимодействия неаллельных генов. Используя конкретные примеры, составьте схемы скрещиваний и охарактеризуйте F1 и F2 при разных типах взаимодействия неаллельных генов.
2.Теория Ф. Жакоба и Ж- Моно о регуляции синтеза и-РНК и белков.
3.У свиней белая щетина (ген «В») доминирует над черной (ген «в»), а наличие сережек (ген «С») — над их отсутствием (ген «с»). Определите генотип белого хряка с сережками, если от спаривания его с черными без сережек свиноматками получено 50% белых поросят с сережками и 50% черных поросят с сережками?
4.Петух с розовидным гребнем (ген «R») спарен с имеющей гороховидный гребень (ген «Р») курицей. От этого спаривания получено 25 потомков с гороховидным гребнем, 24 — с ореховидным, 26 — с розовидным и 22 — с простым (листовидным) -гребнем. Определите генотипы родителей, составьте схему скрещивания и определите генотипы и фенотипы потомков.
5.Какова концентрация доминантного гена «R» (при условии применимости закона Харди-Вайнберга), если гомозиготы по рецессивному гену «r» составляют такой процент от всей популяции: 49, 36, 25, 4? Определите генетическую структуру этих популяций.
1. Основные типы взаимодействия неаллельных генов. Используя конкретные примеры, составьте схемы скрещиваний и охарактеризуйте F1 и F2 при разных типах взаимодействия неаллельных генов
Аллельные гены - гены, определяющие альтернативное развитие одного и того же признака и расположенные в идентичных участках гомологичных хромосом.
Итак, гетерозиготные особи имеют в каждой клетке два гена – А и а, отвечающих за развитие одного и того же признака. Такие парные гены называют аллельными генами или аллелями. Любой диплоидный организм, будь то растение, животное или человек, содержит в каждой клетке два аллеля любого гена. Исключение составляют половые клетки – гаметы. В результате мейоза в каждой гамете остается один комплект гомологичных хромосом, поэтому любая гамета имеет лишь по одному аллельному гену. Аллели одного гена располагаются в одном и том же месте гомологичных хромосом. Схематически гетерозиготная особь обозначается так: А/а.Гомозиготные особи при подобном обозначении выглядят так: А/А или а/а, но их можно записать и как АА и аа. Таким образом, каждый диплоидный организм может иметь не более двух аллелей одного гена, однако в пределах вида число аллелей может быть и существенно больше. В таких случаях говорят о серии множественных аллелей.
Анализирующее скрещивание.
По фенотипу особи далеко не всегда можно определить ее генотип. У самоопыляющихся растений генотип можно определить в следующем поколении. Для видов, использующих другие системы полового размножения, применяют так называемое анализирующее скрещивание. Скрещивание гибридной особи с особью, гомозиготной по рецессивным аллелям, называется анализирующим. При анализирующем скрещивании особь, генотип которой следует определить, скрещивают с особями, гомозиготными по рецессивному гену, т.е. имеющими генотип аа. Анализирующее скрещивание – один из основных методов, позволяющих установить генотип особи, по этой причине оно широко используется в генетике и селекции.
Дигибридное скрещивание при неполном доминировании.
По аналогии с моногибридным скрещиванием ясно, что неполное доминирование по одной или двум парам аллелей может изменить классическое расщепление по фенотипу в f2 – 9:3:3:1 таким образом, что каждому генотипу будет соответствовать определенный фенотип. По решетке Пеннета можно подсчитать, что при дигибридном скрещивании возникает 9 различных генотипов в следующих числовых отношениях. Следовательно, при неполном доминировании по двум парам генов, участвующим в дигибридном скрещивании, следует ожидать 9 фенотипических классов. Именно такое расщепление наблюдается, например, при дигибридном скрещивании у кур, гетерозиготных по гену курчавости оперения и гену, формирующему разбрызганную окраску оперения – черные перышки на белом фоне.
Полигибридное скрещивание.
Понятно, что количество пар генов, по которым могут быть гетерозиготны скрещивающиеся организмы, часто оказываются больше двух. Такое скрещивание называют полигибридным. Количество генотипов и фенотипов, возникающих в таких полигибридных скрещиваниях, резко возрастает, хотя закономерности, которым оно подчиняется, те же, что и в моно- и дигибридном скрещиваниях.
Неаллельные взаимодействия генов
Доминирование и рецессивность – типичные примеры взаимодействия аллельных генов. Однако в процессе индивидуального развития организма и неаллельные гены вступают в сложные взаимодействия между собой. Организм – не мозаика, складывающаяся из действия отдельных и независимых генов, а сложная система последовательных биохимических и морфологических процессов, определяемых совокупностью генов – генотипом
Понятие наследование признака употребляют обычно как образное выражение. В действительности наследуются не признаки, а гены. Признаки формируются в ходе индивидуального развития организма, которые обусловливаются генотипом и влиянием внешней среды.
Принято различать следующие основные типы взаимодействия неаллельных генов:
Комплементарность,
Эпистаз
Полимерия.
Комплементарное взаимодействие генов.
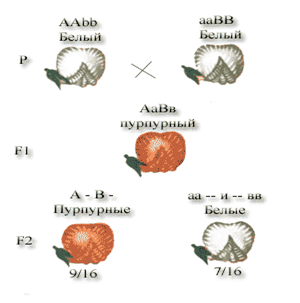
К комплементарным, или дополнительно действующим, генам относятся такие неаллельные гены, которые при совместном проявлении обуславливают развитие нового признака. На примере наследования окраски цветков у душистого горошка можно понять сущность комплементарного действия генов. При скрещивании двух рас этого растения с белыми цветками у гибридов F1 цветки оказались пурпурными. При самоопылении растений из F1 в F2 наблюдалось расщепление растений по окраске цветков в отношении близком к 9:7. пурпурные цветки были обнаружены у 9/16 растений, белые у 7/16. Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментом антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента — пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором -есть пропигмент, но нет фермента, переводящего пропиг-мент в пигмент. Проведем скрещивание двух растений ду шистого горошка с белыми цветками: У дигетерозиготных растений есть и пропигмент (А), и фермент (В), участвующие в образовании пурпурного пигмента. Формирование такого, казалось бы, элементарного признака, как окраска цветков, зависит от взаимодействия по крайней мере двух неаллельных генов, продукты которых взаимно дополняют друг друга. Такая форма взаимодействия генов разных аллельных пар носит название комплементарности — взаимодополнения.
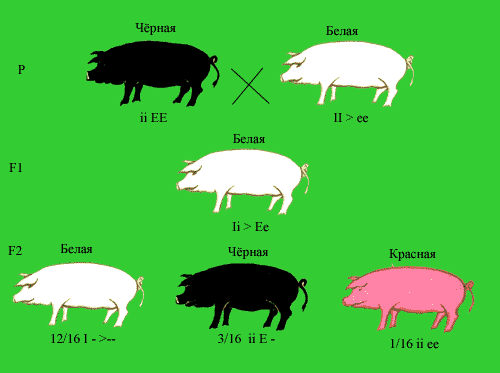
Эпистатическое взаимодействие неаллельных генов в определенном смысле противоположно комплементарному действию генов. Сущность эпистаза сводится к подавлению проявления генов одной аллельной пары генами другой. Гены, подавляющие действие других неаллельных генов, называются супрессорами или подавителями. Они могут быть как доминантными, так и рецессивными, например А - В- или bbA - . Наследование окраски у свиней демонстрирует доминантный Эпистаз. При скрещивании черных и белых свиней из разных пород в F1 появляются белые потомки. Их скрещивание между собой приводит к появлению белых (12/16), черных (3/16) и красных (1/16) поросят. Все белые поросята имеют минимум один доминантный генподавитель I. Черные поросята гомозиготны по рецессивному аллелю i, не препятствующему формированию окраски, и несут доминантный аллель Е, детерминирующий образование черного пигмента. Красные поросята (eeii) лишены доминантного геноподавителя I и доминантного гена, определяющего черную окраску.
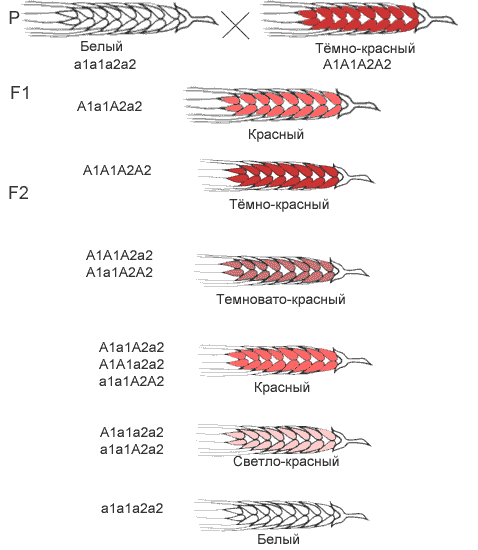
В некоторых случаях установлено, что проявление конкретного признака зависит от количества доминантных генов, вносящих вклад в его развитие. Например, при скрещивании краснозерных пшениц с белозерными было установлено, что растения с генотипом А1А1А2А2 имеют красные зерна, растения а1а1а2а2 – белые зерна, растения с тремя доминантными генами – красноватую окраску, а растения с двумя и одним геном – более бледную окраску. Таким образом, накопление определенных аллелей в генотипе может вести к изменению выраженности признаков.
2. Теория Ф. Жакоба и Ж- Моно о регуляции синтеза и-РНК и белков
Механизм регуляции генетического кода был открыт французскими учеными Ф. Жакобом и Ж. Моно в 1961 г. на бактериях. E. coli и получил название механизма индукции-репрессии. Было установлено, что синтез соответствующих белков – ферментов – индуцируется веществом, служащим субстратом и необходимым для нормальной жизнедеятельности клетки. Так, например, для нормальной жизнедеятельности E. coli необходим молочный сахар (лактоза), и в ее геноме содержаться гены, контролирующие синтез ферментов, гидролизующих лактозу до простых соединений. Если среда, в которой находятся бактерии, лактозы не содержит, эти гены пребывают в репрессированном состоянии и не функционируют. Внесенная в среду лактоза будет тем индуктором, который включает в работу длинные гены, и в клетке начинается синтез ферментов, гидролизующих лактозу до более простых соединений. После удаления лактозы из среды синтез этих ферментов прекращается. Роль репрессора может выполнять и вещество, синтезируемое в клетке, если содержание его превышает норму. Например, если синтезируются нуклеотиды аминокислоты и другие вещества и содержание их превышает количество, необходимое данной клетке, каждое из них может быть репрессором и подавлять работу генов, синтезирующих ферменты, необходимые для данного биохимического процесса.
Механизм индукции-репрессии обеспечивает включение в работу тех генов, которые синтезируют необходимые на данном этапе жизнедеятельности клетки ферменты. Работа генов прекращается, когда деградируемый данными ферментами субстрат израсходован или когда синтезируемое данными ферментами вещество находится в избытке. У высших организмов процесс регуляции работы генов осуществляется более сложно: у животных важную роль в этом процессе играют гормоны, клеточные мембраны; у растений - условия внешней среды, в том числе и окружающие клетки.
Раскрытие механизма регуляции генетического кода показало сложное строение локализованного в молекуле ДНК генетического аппарата. Гены, непосредственно кодирующие синтез соответствующих ферментов, называют структурными генами. Они входят в состав оперона, работу которого регулирует ген-регулятор. Как правило, структурные гены в опероне находятся в состоянии репрессии. Ген-регулятор расположен на особом участке молекулы ДНК и кодирует синтез специального белка, называемого репрессором. Работой структурных генов управляют находящиеся в опероне гены, не имеющие кодирующих функций. Их называют акцепторными генами. Система акцепторных и структурных генов образует единицу генетической регуляции, или оперон.
Акцепторные гены служат местом прикрепления различных белков, регулирующих работу структурных генов. Если лактоза, проникая в клетку, блокирует белки, кодируемые геном-регулятором, то они теряют способность присоединяться к гену-оператору. Ген-оператор переходит в активное состояние и включает в работу структурные гены. РНК-полимераза с помощью Сар-белка присоединяется к промотору и, продвигаясь вдоль оперона, синтезирует про-м РНК. При транскрипции мРНК считывает генетическую информацию со всех трех структурных генов в одном опероне. При трансляции на рибосоме происходит синтез трех разных полипептидных цепей в соответствии с содержащимися в мРНК кодами – последовательностями нуклеотидов, обеспечивающих инициацию и терминацию трансляции каждой цепи.
Тип регуляции работы генов, рассмотренной на примере лактозного оперона, называется негативной индукцией синтеза белка. Другим типом регуляции работы генов служит негативная репрессия, изученная у E.coli на примере trp –оперона, контролирующего синтез аминокислоты триптофана. Этот оперон состоит из 6700 пар нуклеотидов и содержит пять структурных генов, ген-оператор и два промотора. Ген-регулятор обеспечивает постоянный синтез регулярного белка, который не влияет на работу trp-оперона. При избытке в клетке триптофана последний соединяется с регуляторным белком и изменяет его таким образом, что он связывается с опероном и репрессирует синтез соответствующей мРНК.
Известна также и так называемая позитивная индукция, когда белковый продукт гена-регулятора активирует работу оперона, то есть выступает в роли не репрессора, а активатора. Деление это условно, и строение акцепторной части оперона, действие гена-регулятора у прокариотов весьма разнообразны.
Число структурных генов в опероне у прокариотов колеблется от одного до двенадцати; оперон может иметь либо один, либо два промотора и терминатора. Все структурные гены, локализованные в одном опероне, как правило, контролируют систему ферментов, обеспечивающих одну цепь биохимических реакций. Несомненно, что в клетке существуют системы, согласующие регуляцию работы нескольких оперонов.
В связи с особенностями организации отдельных генов эукариот и генома в целом регуляция генной активности у них характеризуется некоторыми отличиями по сравнению с прокариотами.
У эукариот не обнаружено организации генов по типу оперона. Установлено, что функционирование генов, несомненно, подчиняется регуляторным воздействием, однако регуляция транскрипции у эукариот является комбинационной, то есть активность каждого гена регулируется большим спектром генов-регуляторов.
У эукариотических генов имеется несколько областей, которые узнаются разными белками-регуляторами. Одна из них расположена недалеко от промотора и включает около ста пар нуклеотидов, в том числе ТАТА-блок. Установлено, что для успешного присоединения РНК-полимеразы к промотору необходимо предварительное соединение с ТАТА-блоком особого белка – фактора транскрипции – с образованием стабильного транскрипционного комплекса. Именно этот комплекс ДНК с белком узнает РНК-полимеразой. Последовательности нуклеотидов, примыкающих к ТАТА-блоку, формируют требуемый для транскрипции элемент, расположенный перед промотором.
Другая область, играющая важную роль в регуляции генной активности эукариотических генов, располагается на большом расстоянии от промотора и называется энхансером.
И энхансер, и препромоторный элемент эукариотических генов содержат серию коротких нуклеотидных последовательностей, которые связываются с соответствующими регуляторными белками. В результате взаимодействия этих белков происходит включение или выключение генов.
Особенностью регуляции экспрессии эукариотических генов является также существование белко-регуляторов, которые способны контролировать транскрипцию многих генов, кодирующих, возможно, другие белки-регуляторы. В связи с этим белки-регуляторы обладают координирующим влиянием на активность многих генов, и их действие характеризуется плейотропным эффектом. Примером может служить существование белка, который активирует транскрипцию нескольких специфических генов, определяющих дифференцировку предшественников жировых клеток.
Активная работа структурных генов у эукариот зависит от того, какую функцию выполняет клетка в соответствующих тканях или органах. Значительная часть генов в ядрах дифференцированных клеток находится в репрессивном состоянии, при этом регуляция генной активности связанны с образованием стойкого комплекса ДНК с белками – хроматина. Ведущая роль в компактизации ДНК принадлежит гистонам, поэтому они, несомненно, участвуют в процессах регуляции генной активности. Непременным условием для осуществления транскрипции у эукариот является предварительная декомпактизация хроматина на соответствующем участке, где временно утрачивается связь с Н1-гистонами и насколько ослабляется связь с нуклеосомными гистонами. Правда, нуклеосомная организация хроматина не утрачивается даже в ходе транскрипции, однако контакт ДНК и негистоновых белков становится возможным и происходит дерепрессия гена.
Условно структурные гены эукариот могут быть по их активности подразделены на несколько типов. К первому типу могут быть отнесены гены, функционирующие во всех клетках организма. Это гены, кодирующие ферменты энергетического обмена, ферменты, необходимые для синтеза аминокислот, а также гены, контролирующие образование мембраны и других структурных белков. Ко второму типу можно отнести гены, контролирующие синтез миозина в мышечных клетках, коллагена – в костях и т.д. К третьему типу могут быть отнесены гены специализированных клеток, выполняющие важные, но узкие функции – синтез глобина в эритроцитах, гормонов в эндокринных железах и т.д.
Характерно, что в отличие от прокариот мРНК в клетках эукариот может сохраняться довольно продолжительное время, не теряя свои функций. Так, например, у животных ряд типов мРНК синтезируется в оогенезе, сохраняется в яйцеклетке и начинает функционировать на рибосомах после оплодотворения, оказывая значительное влияние на эмбриональное развитие.
Особенностью регуляции транскрипции у эукариот является подчиненность этих процессов регулирующим влиянием со стороны гормонов организма. Последние часто играют роль индукторов транскрипции. Так, некоторые стероидные гормоны обратимо связываются особыми белками-рецепторами, образуя с ними комплексы. Активированный гормоном рецептор приобретает способность соединяться со специфическими участками хроматина, ответственными за регуляцию активности генов, в которых рецепторы узнают определенные последовательности ДНК.
Специфичность регулирующего воздействия гормона на транскрипцию обусловлена не только природой самого гормона, но и природой клетки-мишени, синтезирующей специфический белок-рецептор, который влияет на транскрипцию определенного для данной клетки набора генов. Примером участия гормонов в регуляции активности определенных генов может служить влияние тестостерона на развитие тканей организма по мужскому типу при наличии специфического белка-рецептора. Отсутствие последнего при мутации соответствующего гена не дает возможности гормону проникнуть в ядра клеток-мишеней и обеспечить включение определенного набора генов: развивается синдром тестикулярной феминизации, или синдром Морриса.
У эукариот возможна регуляция синтеза белка на уровне трансляции. При этом имеют значение типы тРНК и ферментов, активирующих соответствующие аминокислоты, а также вырожденность генетического кода. Большая часть аминокислот кодируется несколькими триплетами, получившими название изоакцепторных кодонов. Одна и та же аминокислота может доставляться на мРНК несколькими типами тРНК. Так, кодирование аминокислоты аргинин может происходить посредством кодонов ЦГЦ, ЦГЦ, ЦГА, ЦГГ, АГА, АГГ. Процесс трансляции зависит также от состояния тРНК, рибосом, наличия или отсутствия соответствующих готовые белковые молекулы.
Регуляция на стации трансляции является наиболее экономичной, но недостаточно быстро реагирующей на изменение ситуации. Так, возникшая в клетке потребность в каком-либо белке не может быть быстро удовлетворена путем включения транскрипции соответствующего гена. Синтезированный транскрипт должен подвергнуться процессингу, затем зрела мРНК должна выйти из ядра в цитоплазму и, образуя комплекс с рибосомами, осуществить трансляцию информации, синтезировав пептид, который, лишь пройдя посттрансляционное изменение, формирует активный белок, необходимый клетке.
В том случае, когда клетке нужно прекратить синтез какого-то продукта, после выключения транскрипции соответствующего гена в цитоплазму некоторое время будут продолжать поступать созревающие молекулы мРНК, осуществляющие там синтез пептидных цепей, пока они не деградируют под действием ферментов. Таким образом, для эффективной регуляции экспрессии генов у эукариот должны существовать механизмы, работающие не только на стадии транскрипции, но и на других этапах этого процесса.
Молекула ДНК в процессе биосинтеза осуществляет реализацию наследственной информации. Этот процесс, несмотря на некоторые особенности, характерные для прокариот и эукариот, является, по сути, единым для всего органического мира путем воплощения наследственной информации в свойства и признаки.
3. У свиней белая щетина (ген «В») доминирует над черной (ген «в»), а наличие сережек (ген «С») — над их отсутствием (ген «с»). Определите генотип белого хряка с сережками, если от спаривания его с черными без сережек свиноматками получено 50% белых поросят с сережками и 50% черных поросят с сережками?
Дано:
В-белая щетина
в-черная щетина
С-наличие сережек
с-отсутствие сережок
муж. Ввсс
F1 = 50% В_ С _: 50% ввС_ = 1:1
жен. - ?
Решение:
Р муж. ввсс х жен. ВвСС
F1 ВвСс ввСс
Одну хромосому поток получает от матери, другую от отца, исходя из этого находим генотипы.
Ответ: жен. ВвСС
4. Петух с розовидным гребнем (ген «R») спарен с имеющей гороховидный гребень (ген «Р») курицей. От этого спаривания получено 25 потомков с гороховидным гребнем, 24 — с ореховидным, 26 — с розовидным и 22 — с простым (листовидным) -гребнем. Определите генотипы родителей, составьте схему скрещивания и определите генотипы и фенотипы потомков
Эта задача на признаки, определяющие двумя взаимодействующими генами.
R_P_ - ореховый гребень
rrpp — листовой гребень
R_pp – розовидный гребень
rrP_ – гороховый гребень
F1 по Ф = 25 гор.г. : 24 орех.г. : 26 розов.г. : 22 листов.г. = 1:1:1:1
Р (муж.;жен.) - ?
F1 - ?
Р муж. RrPp x жен. Rrpp
F1 RrPp; rrpp; Rrpp; rrPp
Одну хромосому потомок получает от матери, другую от отца.
Ответ: муж. RrPp; жен. Rrpp
F1 RrPp – ореховый
rrpp – листовидный
Rr pp – розовидный
rrPp – гороховый
5. Какова концентрация доминантного гена «R» (при условии применимости закона Харди-Вайнберга), если гомозиготы по рецессивному гену «r» составляют такой процент от всей популяции: 49, 36, 25, 4? Определите генетическую структуру этих популяций
rr(гомозигота по рецессиву) = 49% = 0,49 (1)
= 36% = 0,36 (2)
= 25% = 0,25 (3)
= 4% = 0,04 (4)
концентрация R -?
(RR + Rr) rr
р + g = 1 з-н Харди-Вайнберга
р — частота встречаемости доминантного гена
р + 0,49 = 1 р + 0,36 = 1 р + 0,25 = 1 р + 0,04 = 1
(RR + Rr) р1 = 0,51 р2 = 0,64 р3 = 0,75 р4 = 0,96
Rr = 2pg — частота гетерозигот
1 Rr = 2 x 0,51 х 0,49 = 0,50 2 2 х 0,36 х 0,64 = 0,46
rr = 0,49 = 49% rr = 36%
(RR + rr) = 0,51 RR = 0,64 — 0,46 = 18%
0,51 — 0,5 = 0,01 Rr = 46%
RR = 0,01 = 1%
Rr = 0,50 = 50%
3 2pg = 2 х 0,75 х 0,25 = 0,38 = 38% 4 2pg = 2 х 0,04 х 0,96 = 0,08
RR = 0,75 — 0,38 = 0,37 = 37% RR = 0,96 — 0,08 = 0,88
Литература:
А.В. Бакай, И.И. Кочиш, Г.Г. Скрипниченко: «Генетика», М. 2006г.