
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
Психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Реферат: Динамическое поведение мембранных систем и липидно-белковые взаимодействия
Реферат: Динамическое поведение мембранных систем и липидно-белковые взаимодействия
Динамическое поведение мембранных систем и липидно-белковые взаимодействия
Введение
Все биологические структуры по своей природе динамичны, и при рассмотрении их функций необходимо учитывать подвижность компонентов, из которых эти структуры состоят. Это относится к ферментам, полинуклеотидам и, конечно, к мембранам. В жидкостно-мозаичной модели, в центре которой находится представление о подвижности мембранных компонентов, мембрана рассматривается как некое липидное море, в котором свободно плавают глобулярные белки. За последние два десятилетия было опубликовано множество работ, посвященных количественным и качественным аспектам динамики мембранных компонентов. Во многих из них использовались спектральные методы и спектроскопия магнитного резонанса. В результате была создана весьма полезная физическая картина мембран, иллюстрирующая способы перемещения мембранных белков и липидов и их взаимодействия. Кроме того, были разработаны специальные методики, направленные на изучение динамических свойств мембран. Многое в этой области остается неясным, но уже определены основные направления будущих исследований.
Главным побудительным мотивом в изучении мембранной динамики служит ее связь с биологическими функциями мембран. Необходимым условием протекания одних ферментативных процессов является свободная диффузия мембраносвязанных компонентов в плоскости бислоя, другие же процессы могут осуществляться лишь при ограниченной подвижности мембранных компонентов. Между шероховатым и гладким эндоплазматическими ретикулумами, комплексом Гольджи и плазматической мембраной происходит быстрый обмен различными веществами, и тем не менее их состав и функции различаются. Чтобы понять суть этих и многих других биологических феноменов, необходимо прежде всего выяснить фундаментальные аспекты динамических свойств мембран. Поперечная асимметрия в распределении липидов, а возможно, и пассивная проницаемость бислоев очевидным образом связаны со скоростью трансмембранного флип-флоп-переноса липидов. Биогенез мембран зависит от скорости обмена липидов между различными мембранами. Скорость ферментативных реакций, протекающих с участием мембраносвязанных компонентов, зависит от скорости латеральной диффузии компонентов мембран. Наконец, липидно-белковые взаимодействия зависят от скорости, с которой происходит обмен липидами между ближайшим окружением белков и остальным объемом мембраны.
Диапазон движений, происходящих в мембране, весьма широк: от молекулярных колебаний с частотой порядка 1014 с до трансмембранного флип-флоп-переноса липидов, характерное время которого может достигать несколько суток. На рис. 1 в общем виде представлены некоторые из этих процессов, а также указаны временные пределы чувствительности различных биофизических методов. Величины, приведенные на рисунке, различаются на 20 порядков, поэтому термины «быстрый» и «медленный» не могут адекватно характеризовать различные типы движений. Из рисунка видно также, что одни методы позволяют получить статичную картину мембраны, поскольку характерное время соответствующих движений больше, чем время измерений, в то время как другие методы дают усредненную по времени картину, поскольку время перемещения молекул гораздо меньше, чем время измерения.
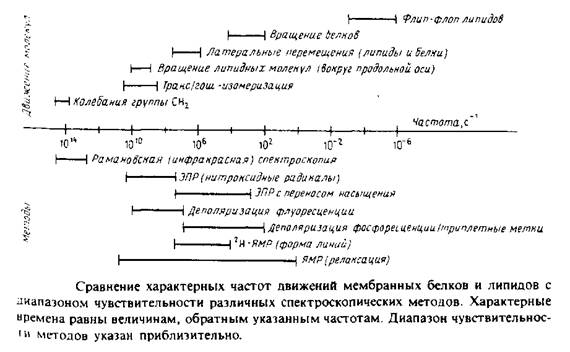
Мы рассмотрим два основных типа экспериментов. Первые основаны на использовании внутримембранных зондов для изучения текучести мембраны. Индикаторами физического состояния мембраны, а также характера липидно-белковых взаимодействий могут служить низкомолекулярные ЭПР-метки и флуоресцентные зонды.
Второй тип экспериментов направлен на прямое измерение латеральной диффузии мембранных белков или липидов и вращательной способности белков внутри бислоя. Исследовались также молекулярные взаимодействия в бислое, поскольку они влияют на динамику изучаемых молекул.
1. Некоторые простые модели движения мембранных компонентов
На рис. 2 представлены некоторые модели, использующиеся для анализа поступательного движения молекул внутри мембранного бислоя. Такие модели необходимы для интерпретации экспериментальных данных с точки зрения молекулярного движения.
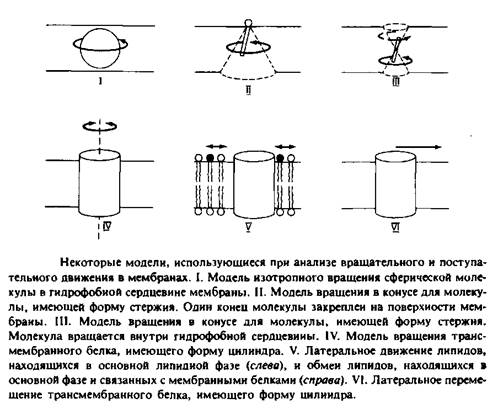
Модель изотропного вращения предполагает, что молекула вращается с одинаковой вероятностью во всех направлениях и выделенная ось вращения отсутствует. Подобное вращение будет совершать сферическая частица в непрерывной жидкой среде. Эта концепция применялась при анализе движения малых растворимых в мембране гидрофобных меток, таких, как TEMPO. Изотропная трехмерная диффузия сферической частицы рассматривается как случайное блуждание и характеризуется коэффициентом вращательной диффузии DBp. Этот параметр определяется как среднеквадратичное угловое отклонение или время релаксации.
2.Модель вращения в конусе описывает движение амфифильных меток, например производных жирных кислот. Эти молекулы можно представить в виде жестких стержней, один конец которых закреплен на поверхности мембраны. Их движение ограничивается конусом, который они описывают относительно некой оси, перпендикулярной плоскости мембраны.
3.Модель вращения в конусе используется в некоторых работах для описания движения ДФГ — гидрофобного флуоресцентного зонда, представляемого в виде жесткого стержня. В данном случае эта модель, постулирующая существование предпочтительной ориентации для ДФГ, является, безусловно, слишком упрощенной, поэтому были предложены альтернативные модели.
4. В простейшей модели вращения белков белковая молекула представляется в виде цилиндра, закрепленного в мембране и способного вращаться только вокруг оси, перпендикулярной плоскости мембраны. Такое вращение анизотропно. Время релаксации, характеризующее это движение, обычно представляется в виде 4>ц = \/. Знак «II» указывает на то, что вращение происходит
параллельно оси цилиндра.
5
и 6. Поступательное движение липидных и белковых
молекул описывается как двумерная диффузия. Особый
интерес представляет скорость обмена липидов, связанных с мембранными белками и
находящихся в основном объеме мембраны. Изотропная двумерная диффузия
характеризуется среднеквадратичным перемещением </2>,
происходящим за время ![]()
Таблица 1. Некоторые метки, использующиеся для изучения динамики мембран
Для обычной жидкости, какой является, например, вода, текучесть определяется как величина, обратная вязкости — понятному и легко измеряемому физическому параметру. Вязкость характеризует трение, возникающее между соседними слоями жидкости, которые движутся с разными скоростями. Вязкость жидкости можно оценить, измерив скорость, с которой падает мраморный шарик в жидкости. В случае мембран термин «текучесть» обычно носит скорее качественный характер: имеется в виду сопротивление, которое оказывает мембрана различным типам перемещений в ней. Как правило, для измерения текучести наблюдают за движением спиновых или флуоресцентных зондов, включенных в мембрану. Зондами обычно являются небольшие молекулы, сравнимые по размерам с мембранными фосфолипидами. Некоторые из них представлены в табл. 5.1. Теоретические основы использования спектроскопических методов для изучения молекулярных движений описаны в последующих разделах. Поскольку эти методы позволяют измерять как скорость движения, так и сопротивление этому движению, сведения о динамике и молекулярной упорядоченности даются вместе. Укажем некоторые моменты, существенные для количественной интерпретации данных по движению зондов внутри мембран.
1.Липидный бислой не является простой вязкой трехмерной гомогенной жидкостью, а представляет собой жидкую среду с низкой вязкостью, у которой состав и динамические свойства в центральной области сильно отличаются от состава и свойств периферических полярных участков.
2.Вращение зондов изотропно, как это имеет место в случае сферических частиц, не обладающих выделенной осью вращения. Часто зонды внутри мембраны имеют предпочтительную ориентацию и их движения ограничены определенными рамками. Интерпретация экспериментальных данных зависит от модели, используемой для описания молекулярного движения.
3.Локализация зондов в мембране может различаться. Например, зонд может быть связан с белковой молекулой или белковыми агрегатами, располагаться внутри липидного домена, который может находиться в различных физических состояниях.
Для оценки текучести определяют спектроскопические параметры низкомолекулярных зондов. К таким параметрам относятся: 1) время вращательной корреляции спиновых меток или флуоресцентных зондов; 2) параметр упорядоченности; 3) стационарная анизотропия флуоресцентных зондов; 4) коэффициент распределения зондов между мембранной и водной фазами. Ясно, что текучесть мембраны, оцененная только по одному параметру, не может служить достаточно полной характеристикой физического состояния мембраны. И все же измерение отдельных параметров весьма полезно, особенно для характеристики изменений физического состояния мембраны, обусловленных, например, изменениями температуры, давления, содержания холестерола, фосфолипидного или ионного состава. Обычно текучесть, измеряемая с помощью спиновых меток и флуоресцентных зондов, однозначно связана с упаковкой липидов в мембране. Воздействия, приводящие к уменьшению площади, приходящейся на одну липидную молекулу, такие, как увеличение гидростатического давления, понижение температуры или добавление холестерола к фосфолипидам в жидкокристаллическом состоянии, вызывают уменьшение текучести. Это согласуется с теорией свободного объема, согласно которой текучесть и плотность связаны между собой обратной зависимостью. Чем более плотная упаковка характерна для мембраны, тем более ограниченным будет движение зонда. Такой подход приемлем при рассмотрении текучести большинства неассоциированных жидкостей.
2. Физиологическое значение текучести мембран
Обычно биомембраны находятся в жидкокристаллическом состоянии, и, по-видимому, поддержание такого состояния очень важно для их функционирования. При переходе мембраны из жидкокристаллической фазы в фазу геля текучесть уменьшается примерно на два порядка. Структурные и динамические свойства бислоя, находящегося в фазе геля, совершенно несовместимы с организацией и правильным функционированием белковых компонентов в мембране. Впрочем, из этого правила имеются несколько исключений. Это, например, полукристаллические области пурпурных мембран Н. halobium, содержащие бактериородоп-син.
Возможно, наиболее яркое доказательство того, что текучесть, измеряемая с помощью спиновых меток или флуоресцентных зондов, играет важную физиологическую роль, получено в исследованиях по адаптации различных организмов к внешним экстремальным воздействиям. Подобные явления наблюдаются чаще всего при изучении термического стресса, когда микроорганизмы, растения, пойкилотермные или зимующие животные подвергаются воздействию низких температур. Адаптация заключается в изменении липидного состава мембран, а именно — в увеличении содержания ненасыщенных липидов или уменьшении средней длины ацильной цепи. Подобные изменения ведут к уменьшению плотности упаковки липидов в мембране и, таким образом, поддерживают текучесть мембраны. Текучесть мембран может быть критичной для одной или более мембранных функций, но каков механизм этого феномена на молекулярном уровне — неизвестно.
2.1 Характер и скорость движений, измеряемых с помощью 2Н-ЯМР, ЭПР и флуоресцентных зондов
Необходимо иметь представление о том, что именно измеряется с помощью мембранных зондов; особенно важно знать, каково отношение между временным интервалом, в котором наблюдается изучаемый тип движения, и временной шкалой, характерной для данного метода. Рассмотрим общий случай, который одинаково применим к ЭПР, измерению флуоресценции или 2Н-ЯМР. Все три спектроскопических метода чувствительны к ориентации молекул относительно лабораторной системы координат. Спектр 2Н-ЯМР чувствителен к ориентации связи С—D относительно приложенного поля. Это связано с тем, что градиент локального поля, векторно суммируясь с внешним полем, дает результирующую, которая и улавливается дейтерием. Аналогичная ситуация характерна и для ЭПР. Спектр ЭПР зависит от ориентации нитроксидной связи N— О, которую содержат большинство обычно используемых зондов, относительно приложенного магнитного поля. В случае флуоресцентной спектроскопии измеряемая поляризация испускаемого света зависит от ориентации дипольного момента перехода молекулы относительно направления, определяемого используемым поляризатором.
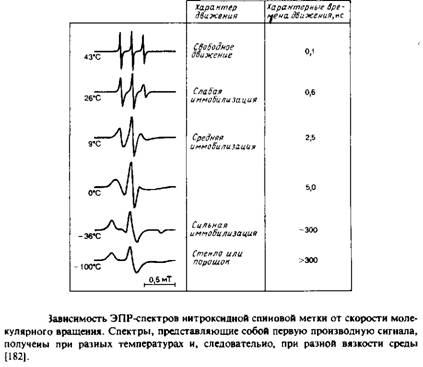
Если молекулы образца ориентированы случайным образом и полностью неподвижны, то каждая субпопуляция, характеризующаяся определенной ориентацией, будет иметь свой уникальный спектр ЭПР или ЯМР, а спектр всего образца будет равен простой сумме этих спектров с учетом их вкладов, определяемых концентрацией соответствующих молекул. Суммарный спектр называется «порошковым» или «твердостекольным» и обычно весьма широк. Такой спектр сильно отличается от спектра образца, в котором молекулы также ориентированы случайным образом, но характерное время их движений гораздо меньше времени измерения. В рассматриваемых нами примерах соответствующие частоты составляют 105 с-1 для 2Н-ЯМР и 108 с"1 для ЭПР. Каждая молекула, совершающая быстрое изотропное движение, воспринимает некое среднее магнитное поле, но при этом для всех молекул это среднее поле одинаково, поэтому пики в результирующем спектре очень острые. Примеры ЭПР-спектров приведены на рис. 5.3. Поскольку характерные времена измерения для методов ЭПР и ЯМР различаются, молекула со временем корреляции 106 с будет казаться неподвижной в первом случае и очень быстро движущейся во втором.
Метод измерения деполяризации флуоресценции состоит в том, что образец облучают поляризованным светом и спустя несколько наносекунд регистрируют испускаемый свет. Если молекулы остаются неподвижными в течение времени между процессами поглощения света и его испускания, то поляризация испускаемого света будет определяться в основном поляризацией света возбуждающего. Если же молекула совершает быстрое изотропное вращение, то вектор напряженности электрического поля испускаемого света не будет иметь предпочтительной ориентации, поскольку поляризация будет равна нулю. Значения поляризации, находящиеся между этими двумя крайними величинами, соответствуют или медленным движениям, при которых популяция молекул не успевает достичь полностью разупорядоченного состояния за время 10-8 с, или быстрым движениям, ограниченным в пространстве и, следовательно, не допускающим полную разупорядоченность.
Все три спектроскопических метода используются для нахождения параметра упорядоченности S:
![]()
cos20 — это усредненная по времени ориентация молекулы относительно нормали к плоскости мембраны. Время, за которое происходит усреднение, зависит от метода: 10~5 с для 2Н-ЯМР, 10~8 с для ЭПР и деполяризации флуоресценции. В случае 2Н-ЯМР этот параметр чувствителен к медленным движениям, совершающимся за времена порядка 10"5 с. Метод ЭПР или деполяризации флуоресценции регистрирует процессы, совершающиеся за время около 10"8 с, а информация о более медленных движениях утрачивается.
Данные о динамических свойствах мембран можно получить с помощью всех трех методов и более прямым способом. Так, из данных ЯМР можно найти времена релаксации, которые в свою очередь зависят от характера молекулярных движений. Из спектра ЭПР можно при определенных условиях найти время вращательной корреляции зонда. Измеряя зависимость степени деполяризации флуоресценции от времени, можно получить данные о динамическом состоянии зонда, а не только оценить степень ограниченности его движения.
2.2 Спиновые метки, использующиеся при изучении текучести мембран
ЭПР — это очень ценный метод изучения мембран. Обычно в качестве меток используются стабильные парамагнитные соединения, содержащие нитроксидный радикал. Неспаренный электрон при наложении сильного магнитного поля переходит с одного энергетического уровня на другой под действием микроволнового излучения. Метод очень чувствителен: обычно спектр регистрируется при концентрации спиновых меток около 10"*М в 50 мкл образца. Спектр ЭПР, как правило, представляют в виде первой производной от спектра поглощения.
В спектре нитроксидного радикала имеются три пика, отвечающие спин-спиновым взаимодействиям неспаренного электрона и ядра атома азота. И положение спектра, и расщепление, обусловленное спином ядра, зависят от ориентации молекулы относительно внешнего поля. Как мы уже говорили в предыдущем разделе, это означает, что характер спектра зависит от характера вращения молекулы. На рис. 5.3 приведены спектры для самых разных случаев — от свободного вращения до полной неподвижности. Для того чтобы найти тк в предположении, что вращательное движение молекулы изотропно, можно использовать простое уравнение, в которое входит высота пиков. Этот подход наиболее применим для такой метки, как TEMPO.
Спиновые метки — производные жирных кислот или фосфолипидов, — конечно, не вращаются изотропно, и в этом случае для нахождения параметра упорядоченности S из величины расщепления между линиями спектра используется вторая простая формула. Предположив, что такие метки имеют форму жесткого стержня, можно из величины параметра S оценить максимальное отклонение зонда от нормали к поверхности мембраны.
Многие предположения, которые делаются при нахождении из спектров ЭПР времен вращательной корреляции или параметра S, снижают ценность этого метода для определения количественных характеристик молекул. Эти параметры тем не менее позволяют получить качественное представление о поведении мембраны. Результаты измерения степени упорядоченности спиновых меток, фиксированных в мембране на разной глубине, свидетельствуют об увеличении неупорядоченности в направлении от поверхности мембраны к ее центральной области. Качественно такая же картина получена с помощью 2Н-ЯМР. Во многих исследованиях проводилось сравнение текучести мембран, которую определяли по данным о величинах тк и S для спиновых меток, при различных возмущающих воздействиях.
2.3 Флуоресцентные зонды, используемые для измерения текучести мембран
Для измерения вращательной диффузии молекул давно используется метод деполяризации флуоресценции. Вращение некоторых зондов в мембранах часто сравнивают с их вращением в маслах с известной вязкостью, при этом пользуются понятием «микровязкость», которую измеряют в пуазах. Термин «микровязкость» означает, что зонд «чувствует» величину вязкости только в своем непосредственном окружении, т. е. этот параметр не является макроскопическим. С точки зрения молекулярного движения подобные измерения не имеют строгого количественного смысла, но, как и в случае со спиновыми метками, полученные результаты позволяют создать ценную качественную картину.
В табл. 1 приведены структурные формулы некоторых зондов, используемых при измерении текучести мембран. Чаще всего применяют ДФГ, поскольку он с легкостью накапливается в мембранах, обладает интенсивной флуоресценцией, по-видимому, не связывается с белками и чувствителен к физическому состоянию мембран. Однако временные измерения свидетельствуют о некой гетерогенности неизвестной природы; кроме того, имеются разногласия по поводу ориентации зонда в бислое и характера его подвижности.
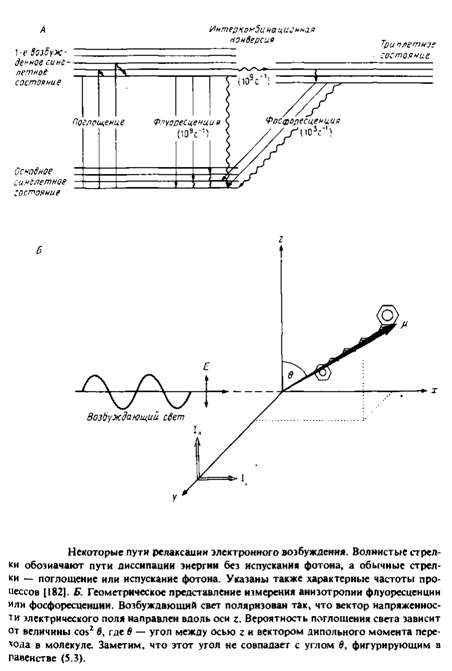
Рис. 4 иллюстрирует принципы, лежащие в основе применения флуоресценции и фосфоресценции для анализа вращения молекул. Молекула, поглотившая фотон, за время 10" 11 с переходит в возбужденное синглетное состояние Si. Термин «синглетное» указывает на то, что спин электрона при поглощении фотона не изменяется. Обычно возвращение в основное, невозбужденное состояние происходит за время - 10"8 с. Причинами такого перехода являются столкновение с соседними молекулами, диссипация энергии в виде тепла или испускание фотона. Характерное время перехода популяции молекул из синглетного возбужденного состояния в основное равно тт. Некоторые молекулы с большой вероятностью переходят из синглетного возбужденного состояния в триплетное, и этот переход сопровождается изменением спина электрона. Испускание фотона при переходе молекулы из этого состояния в основное называется фосфоресценцией. Время жизни молекулы в триплетном состоянии относительно велико, что связано с изменением спинового состояния электрона при испускании фотона.
Рассмотрим принцип метода деполяризации. Свет, поляризованный вдоль оси z, будет предпочтительно возбуждать молекулы, у которых дипольный момент перехода имеет такую же ориентацию. Вероятность возбуждения пропорциональна cos2 в, где в — угол между дипольиым моментом перехода и вектором напряженности электрического поля. Такая же зависимость от cos2 в имеет место и для процесса испускания фотона, поэтому молекула, у которой дипольный момент перехода ориентирован вдоль оси z, будет испускать свет с такой же поляризацией.
Испускаемый свет анализируют с помощью поляризаторов, определяя интенсивность компоненты, параллельной и перпендикулярной направлению поляризации возбуждающего света.
Анизотропия г связана с /, и IL следующим образом:
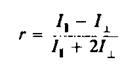
Если между моментами поглощения и испускания света молекула не вращается, то /, будет больше, чем 1±, и г будет иметь максимальное значение 0.4. Если же молекулы, находясь в возбужденном состоянии, совершают интенсивные изотропные вращательные движения, то г будет равно нулю, поскольку в среднем молекулы к моменту испускания света будут ориентированы беспорядочно. При тк = tf величина г будет зависеть от характера движения. При этом т ~ 1 и ту ' отражают скорости вращения и испускания света.
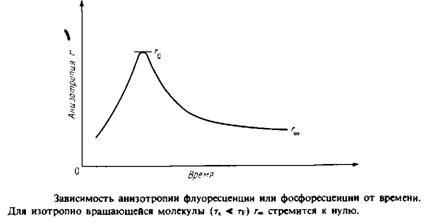
Для того чтобы разграничить динамические эффекты и эффекты, связанные с ограничениями, налагаемыми на диапазон допустимых движений, определяют временную зависимость анизотропии r света, испускаемого молекулой после возбуждения. Значение г в данный момент времени отражает степень разу-порядочивания молекул относительно вектора поляризации за вре- ■ мя, прошедшее после поглощения возбуждающего импульса света. Спад r от максимального значения го задается выражением
![]()
тк — время вращательной корреляции, оно определяется скоростью затухания. Если при / оо все молекулы окажутся беспорядочно ориентированными, то л в пределе будет равно нулю. Однако если движение молекул ограничено, то их ориентация никогда не будет совершенно случайной. В такой ситуации г» не будет равно нулю, и с помощью этого параметра можно определить степень ограничения, которое налагается на подвижность зонда его окружением. Параметр упорядоченности определяется из этих величин так же, как в случае ЭПР или ЯМР:
![]()
К сожалению, такие кинетические измерения проводятся редко, большинство экспериментов выполнено в стационарном режиме при непрерывном возбуждении и испускании. Значение г, полученное таким способом, носит усредненный характер и определяется как
![]()
Рассмотрим следующие три случая.
1. Очень быстрое движение: 7"= г„, предельному значению.
2. Очень медленное движение или его отсутствие: г = го, максимальному значению.
3. Случаи, представляющие практический интерес '- 7 зависит как от скорости движений, так и от ограничений, налагаемых на них.
Без соответствующей корректировки определение микровязкости по результатам измерений 7 не будет сводиться к простым динамическим измерениям.
Вязкость бислоя весьма высока, поэтому время вращательной корреляции небольших флуоресцентных и ЭПР-зондов составляет 10~8—10 с. В воде, имеющей вязкость 0,01 пуаз, молекулы такого размера вращались бы по крайней мере в 100 раз быстрее. Мембранные белки имеют значительно большие размеры, чем упомянутые метки, и поэтому вращаются гораздо медленнее. Чтобы метки, «пришитые» к этим белкам, были чувствительны к их вращению, время жизни меток в возбужденном состоянии должно составлять порядка 10~3 с.
3. Вращение мембранных белков
Коэффициент вращательной диффузии мембранного белка, находящегося в плоскости бислоя, D., можно найти, представив белко
вую молекулу в виде цилиндра, который вращается вокрут одной оси. Пусть мембрана имеет вязкость г) и толщину h, а радиус цилиндра равен а. Тогда
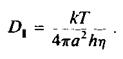
Часто наряду с фу применяется время вращательной релаксации ф, = 1/. Для белковой молекулы радиусом 25 А, находящейся в мембране толщиной 40 А и вязкостью 5 пуаз, величина ф, по оценкам составляет около 35 мкс. С количественной точки зрения это уравнение, описывающее вращение белка в бислое, не вполне строго, но зависимость времени вращательной релаксации от эффективного радиуса вращающейся белковой молекулы сомнений не вызывает. Это оказалось весьма полезным для исследования процессов агрегации белков внутри мембраны. Методы, применяемые для изучения вращения белков в бислое, должны быть способны регистрировать времена вращения от 10"5 до 10 ~3 с. Обычный метод измерения деполяризации флуоресценции в этом случае непригоден, поскольку время жизни молекул в возбужденном состоянии составляет около 10"8 с, и в таком временном масштабе молекулы белков представляются неподвижными. Успешно использовались три метода.
1. К исследуемому белку присоединяют зонд, время жизни которого в возбужденном триплетном состоянии достаточно велико. Если метка жестко связана с белком, то для регистрации вращения белка можно использовать измерение анизотропии фосфоресценции. Для таких измерений оказались пригодными производные эозина, поскольку время жизни эозина в триплетном состоянии составляет примерно 2 мс. В основе этого подхода лежат те же принципы, что и в основе метода деполяризации флуоресценции, но используется другая молекулярная модель движения, вызывающего деполяризацию. Эксперимент состоит в определении характерного времени затухания анизотропии фосфоресценции. Если система гетерогенна, могут возникнуть определенные трудности, связанные с количественным анализом экспоненциальной кривой затухания анизотропии. Проблемы возникают и в тех случаях, если метка может локально вращаться на поверхности мембранного белка или если у белковой молекулы имеются гибкие сегменты.
Те же метки могут использоваться и для анализа вращения белков с использованием временной зависимости дихроизма поглощения. В этом случае изменения в ориентации дипольного момента перехода регистрируются по поглощению света, поляризованного параллельно и перпендикулярно оси поляризации начального импульса, используемого для возбуждения молекул.
2. Известны случаи, когда сами молекулы белка содержат группы, переходящие при флеш-фотолизе в долгоживущее возбужденное состояние, параметры которого можно оценить с помощью дихроизма поглощения. В качестве примера можно привести родопсин и бактериородопсин, где используются возбужденное состояние связанного ретиналя и возбужденные состояния, наблюдающиеся при фотолизе комплексов цитохром—СО с использованием цитохром с-оксидазы и цитохрома Р450. Измерения можно проводить in situ или с очищенным белком, встроенным в фосфолипидные везикулы.
3. С помощью обычной ЭПР-спектроскопии не удается регистрировать вращения, характерная частота которых равна частоте вращения мембранных белков. Однако разработан специальный метод — ЭПР с переносом насыщения, диапазон чувствительности которого очень широк — от 10"7 до 10 ~J с. Этот метод применялся при изучении вращения нескольких мембранных белков с ковалентно пришитыми к ним спиновыми метками. Недостаток метода состоит в том, что в случае анизотропного молекулярного движения спектры с трудом поддаются интерпретации.
3.1 Примеры вращения белков
Внутримембранные белки характеризуются широким спектром времен вращательной релаксации. На одном конце временной шкалы находится родопсин, который, по-видимому, свободно вращается в мембране наружного сегмента палочки сетчатки, а на другом — бактериородописин, который образует в пурпурной мембране упорядоченную кристаллическую решетку и неподвижен. Для нескольких очищенных белков, встроенных в фосфолипидные везикулы, зависит от концентрации. Это позволяет предположить, что при уменьшении соотношения липид/белок происходит самоагрегация этих белков. Такая картина характерна для бактериородопсина, цитохром с-оксидазы, белка полосы 3, Са2 + -АТРазы и цитохрома Р450. Возможно, именно с самоагрегацией связана гетерогенность, наблюдаемая, в частности, для белка полосы 3 в тенях эритроцитов и Са2 + -АТРазы из саркоплазматического ретикулума. Вообще говоря, мембранные белки, по-видимому, вращаются в плоскости мембраны и скорость вращения согласуется с величиной, ожидаемой исходя из простой гидродинамической модели.
Дополнение. Вращение белка полосы 3 и цитохрома Р450: выявление межмолекулярных взаимодействий
Из двух указанных белков — белка полосы 3 и цитохрома Р450 — наибольший интерес представляет первый, поскольку по имеющимся биохимическим данным можно предположить, что этот анионный переносчик связан со спектрино-актиновой сетью цитоскелета. Обнаружение того факта, что значительная часть белковых молекул способна к быстрому вращению, послужило основанием для создания моделей, согласно которым белок полосы 3 иммобилизован не полностью или образует со спектрином временную связь. Однако ситуация осложнялась тем, что при лизисе клеток, необходимом для получения теней эритроцитов, комплекс с цитоскелетом может разрушаться. В интактной клетке белок полосы 3 совершает только медленные вращательные движения с характерным временем 0,1 — 1 мс. Это больше согласуется с отсутствием поступательного движения белка полосы 3, хотя физическая причина такого ограничения подвижности не установлена.
Цитохром Р450 акцептирует электроны от цитохром Р450-редук-тазы в электронтранспортной цепи микросом. Многих ученых интересовал вопрос: образуют ли эти молекулы долгоживущий комплекс в мембране или их взаимодействие сводится к простым столкновениям в результате диффузии? При низком соотношении липид/белок цитохром Р450 агрегирует и вращение его замедляется. Однако в присутствии стехиометрических количеств редуктазы цитохром Р450 в сходных условиях остается мобильным. Это свидетельствует о взаимодействии двух указанных белков и образовании комплекса между ними со стехиометрией 1:1, которое препятствует агрегации Р450.
4. Латеральная диффузия липидов и белков в мембранах
Способность мембранных компонентов к латеральной диффузии — это одно из очевидных условий, принятых в жидкостно-мозаичной модели. Для измерения коэффициента латеральной диффузии применялось несколько методов. Рассмотрим три из них.
1.Осуществляют слияние клеток, несущих на своей поверхности различные маркеры, с образованием гетерокарионов. После слияния оценивают скорость перераспределения поверхностных маркеров с помощью флуоресцентной микроскопии и с использованием меченных различными флуоресцентными метками антител против специфических поверхностных антигенов.
2.Осуществляют перераспределение мембранных белков с помощью электрофореза in situ. При внесении клеток в электрическое поле белки концентрируются в одной части мембраны, и коэффициент латеральной диффузии определяют по данным о скорости их перераспределения после отключения электрического поля. За перераспределением белков можно наблюдать с помощью электронной микроскопии или флуоресцентных методов.
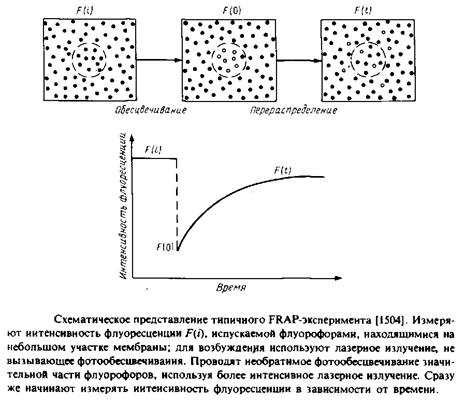
3.Чаще всего используется метод, основанный на восстановлении способности белков к флуоресценции после фотообесцвечивания. Суть метода представлена на рис. 5.6. С помощью мощного лазерного луча обесцвечивают пятно диаметром - 1 мкм на равномерно меченной с помощью флуоресцентной метки мембране. Далее исследуют кинетику разгорания флуоресценции этого участка и по скорости восстановления флуоресценции прямо оценивают скорость латеральной диффузии флуоресцентных зондов на обесцвеченный участок из областей мембраны, прилегающих к нему. С помощью этого метода можно исследовать амфифильные флуоресцентные зонды, меченые фосфолипиды и белки, несущие флуоресцентные метки. Структурные формулы некоторых наиболее часто используемых зондов приведены в табл. 5.1. Можно ожидать, что применение видеотехники для изучения перераспределения флуоресцентных молекул в мембране расширит рамки применения этого метода. Использование двух первых методов сопровождается значительными нарушениями клеток или мембран. Метод FRAP является более щадящим. Опасения, связанные с возможностью повреждений клеток лазерным лучом, были сняты. Метод можно также применять для исследования модельных мембранных бислоев и монослоев, а также интактных клеток или фрагментов биомембран. Фокусируя лазерный луч диаметром ~ 1 мкм в разных точках, можно наблюдать за датеральной диффузией мембранных компонентов в различных участках мембраны.
Коэффициенты поступательной диффузии, которые можно определить с помощью этого метода, варьируют от Ю-7 до 10 ~12 см2/с. Коэффициент диффузии 10 "12 см2/с в рамках этого подхода соответствует отсутствию движения. Значение D = 10 "8 см2/с, характерное для липидов в биомембранах, отвечает диффузии на расстояние около 2 мкм за 1 с.
4.1 Модели, описывающие латеральную диффузию
Для описания поступательной диффузии мембранных белков часто применяется гидродинамическая модель Саффмана и Дельбрюка. В этой модели рассматривается диффузия белка в тонком вязком слое; при этом считается, что растворитель является сплошной средой, т. е. молекулы растворителя малы по сравнению с диффундирующими молекулами. В рамках этой модели выполняется следующее соотношение:
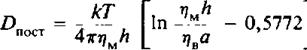
где т)м и 1), — вязкости мембранной и водной фаз соответственно, а — радиус белковой молекулы, имеющей форму цилиндра, h — толщина мембраны. Из модели следует, что скорость поступательной диффузии слабо зависит от размера молекулы, что подтверждается полученными к настоящему времени экспериментальными данными.

Эта гидродинамическая модель не позволяет описывать диффузию
липидных зондов, поскольку к ним неприменимо допущение о том, что белок
диффундирует в сплошной среде. Для объяснения поведения липидов применялись
различные модели, основанные на концепции свободного объема. В рамках этих
моделей диффузия рассматривается как процесс движения молекул в полостях,
образующихся вследствие спонтанных флуктуации, при этом определяющей характеристикой
мембран является отношение площади, приходящейся на одну молекулу липида, к
минимальной Удельной площади, соответствующей максимально плотной упаковке
липидов, т. е. плотность упаковки липидов. Как мы уже отмечали, скорость
вращения флуоресцентных зондов зависит от плотности упаковки липидов в
мембране. Применимость этой модели к описанию вращательной или поступательной
диффузии липидов строго не доказана, поскольку изначально она была построена
для неассоциированных жидкостей, к которым можно отнести далеко не все
мембранные липидные структуры. Тем не менее обратная зависимость между
скоростью поступательной диффузии липидов и плотностью упаковки липидов в
мембранном бислое обычно существует.
Наконец, несколько подходов использовалось для теоретического объяснения влияния мембранных белков на латеральную диффузию белков и липидов в мембране. Дело в том, что белки оказываются на пути диффундирующих молекул и вынуждают их перемещаться только в пространстве, не занятом белками. Обычно из-за этого коэффициенты поступательной диффузии в биомембранах оказываются примерно в 10 раз меньше, чем в чисто липидных бислоях.
4.2 Примеры латеральной диффузии компонентов мембран
Коэффициенты поступательной диффузии некоторых мембранных липидов и белков приведены в табл. 5.2. Коэффициенты диффузии флуоресцентных меток и аналогов липидов в фосфолипидных мультибислоях или в крупных однослойных везикулах равен 10"8 см2/с. При изучении поведения этих же зондов в разнообразных природных мембранах коэффициент диффузии обычно оказывается примерно в 10 раз меньше. Этот феномен, как правило, объясняют присутствием в биомембранах белков, препятствующих латеральной диффузии. Скорость латеральной диффузии фосфолипидов с разными полярными головками различается слабо, однако в реконструированных везикулах гангли-озиды диффундируют медленнее. При переходе мембранного бислоя в состояние геля скорость латеральной диффузии уменьшается более чем на два порядка {D < 10" 10 см2/с). В жидком бислое модельных мембран и в биомембранах липиды обычно диффундируют свободно, хотя имеются редкие исключения.
Совсем иначе обстоит дело с латеральной диффузией мембранных белков. Исследование поведения нескольких белков в реконструированных везикулах, пока не позволяющее сделать окончательные выводы, тем не менее указывает на то, что коэффициенты поступательной диффузии белков разных размеров очень близки и что белки диффундируют лишь немного медленее, чем липиды. Исследованные белки сильно различались по числу трансмембранных сегментов и по степени ассоциированности, и тем не менее диффундировали с одинаковой скоростью. Модель Саффмана—Дельбрюка предсказывает очень слабую зависимость скорости диффузии от размеров молекул. В реконструированных везикулах с высоким содержанием белка могут происходить агрегация последнего и латеральное разделение фаз, приводящие к иммобилизации части белка.
В биомембранах коэффициенты латеральной диффузии белков обычно в 100—1000 раз меньше, чем в модельных системах с низкой концентрацией белка. Например, родопсин, по-видимому, свободно диффундирует в биомембране— D = 4-10"9 см2/с. Поскольку мембраны, содержащие родопсин, характеризуются высоким отношением белок/липид, ясно, что обычно наблюдаемая медленная диффузия других мембранных белков не может быть обусловлена исключительно присутствием белков в бислое. Есть и еще один интересный факт: восстановление флуоресценции в экспериментах FRAP происходит не полностью и нередко составляет менее 75%. Это означает, что часть изучаемой популяции белков ^ неподвижна.
Вопрос о том, что ограничивает латеральную диффузию мембранных белков, имеет ключевое значение. Маловероятно, что это обусловлено агрегацией мембранных белков, поскольку для объяснения большинства данных потребовалось бы предположить, что образуются слишком крупные агрегаты. Одно из возможных объяснений состоит в том, что мембранные белки,малоподвижны, поскольку связаны с внеклеточным матриксом или с ци-тоскелетом. Обнаружилось, что, оказывая воздействие на внеклеточный матрикс, можно влиять на латеральную диффузию мембранных белков, однако имеются и другие данные, согласно которым эта связь не столь уж важна.
О взаимодействиях между мембранными белками и элементами цитоскелета, ответственных за некоторые из наблюдаемых ограничений латеральной диффузии белков, свидетельствуют разные данные. Белок полосы 3 в мембранах эритроцитов в норме в основном неподвижен, но в клетках с недостатком спектрина его латеральная подвижность возрастает по меньшей мере в 40 раз. При нарушениях цитоскелета в тенях эритроцитов латеральная диффузия белка полосы 3 также возрастает. Проводились опыты по изучению диффузии ацетилхолиновых рецепторов во вздутиях в мембранах, где связь с актином и элементами цитоскелета нарушена. Подвижность белка в таких системах была гораздо выше, чем в клетках с интактным цитоскелетом. Тем не менее, гипотезу о том, что низкая подвижность мембранных белков обусловлена их связью с цитоскелетом, нельзя считать бесспорной.
Известен по крайней мере один случай, когда взаимодействия с цитоскелетом не сказываются на латеральной диффузии. G-белок вируса везикулярного стоматита находится в плазматической мембране инфицированных животных клеток и имеет единственный трансмембранный сегмент. Были получены мутантные формы этого белка, у которых цитоплазматический домен отсутствовал. Ни для одного из мутантов не наблюдалась быстрая латеральная диффузия, которая характерна для мембранных белков, встроенных в искусственные бислои. Эти результаты свидетельствуют о том, что прямые взаимодействия между данным белком и цитоплазматическими белками не приводят к уменьшению латеральной подвижности.
Наконец, следует отметить, что с помощью метода FRAP, используемого в большинстве этих экспериментов, была зарегистрирована диффузия на расстояние от нескольких сотен А до 1 мкм. Белок может свободно диффундировать внутри небольшого домена, но встречать препятствия на своем пути в присутствии других интегральных или периферических белков, и при использовании метода FRAP будет считаться неподвижным. Кроме того, если бы взаимодействие между мембранными белками и белками цитоскелета было относительно слабым и частота их ассоциации и диссоциации была достаточно высока, белок мог бы перепрыгивать с одного сайта связывания на другой. В результате суммарная скорость латеральной диффузии уменьшилась бы, поскольку белок большую часть времени находился бы в связанном с цитоскелетом состоянии, но скорость движения белка между сайтами связывания была бы высока. Образование кластеров мембранных белков за счет притяжения между ними также приводит к уменьшению латеральной диффузии.
5. Липидно-белковые взаимодействия
Большинство методов, применяемых для изучения упорядоченности и динамических свойств мембран, используется и для исследования липидно-белковых взаимодействий. Работы по изучению этих взаимодействий были в основном направлены на выяснение влияния мембранных белков на физическое состояние липидов. Рассмотрим типичную мембрану с весовым соотношением липид/белок = 1:1. При средней мол. массе белка 50 кДа молярное соотношение липид/белок составляет -60:1 при условии, что присутствуют только фосфолипиды. Для сравнения укажем, что соответствующие соотношения для мембраны наружного сегмента палочки сетчатки и саркоплазматического ретикулума составляют по оценкам 75:1 и 110:1. Если белок представляет собой цилиндр, выступающий за пределы бислоя с обеих сторон примерно на 10 А, то его радиус должен составлять около 18 А. Молекула фосфолипида в жидкой мембране занимает площадь ~60А2, что соответствует радиусу головки ~4,4 А, и для того, чтобы полностью окружить белок, иа каждой стороне бислоя должно находиться около 16 таких молекул липидов. Следовательно, согласно этой модели, в любой момент времени около 50% липидов должно соседствовать с белковыми молекулами. Однако белком занято 35% площади поверхности мембраны, и даже учитывая сугубо приближенный характер этой модели, можно понять, что физические методы регистрации состояния липидов должны учитывать влияние белков на свойства биомембран.
Чтобы выяснить структуру и функции мембран, необходимо прежде всего ответить на следующие вопросы: 1) насколько прочно связаны внутримембранные белки с липидами и какова природа ли-пидного слоя, прилегающего к белку? 2) как далеко простирается влияние мембранных белков на укладку и динамические свойства мембранных липидов? 3) как влияют липиды на структуру и функции внутримембранных белков? 4) как периферические мембранные белки, связанные с поверхностью бислоя, взаимодействуют с липидами и влияют на их поведение?
5.1 Связывание липидов с внутримембранными белками в бислое
Этой теме были посвящены многочисленные исследования, в которых использовались разнообразные подходы. По существу во всех этих работах ставилась задача выяснить, есть ли у белков участки, специфически взаимодействующие с определенными липидами, и можно ли считать белково-липидные комплексы долгоживущими, т. е. обладают ли они временем жизни, сравнимым с временем оборота типичного фермента. Такие исследования проводились с помощью Н-ЯМР, ЭПР и флуоресцентных методов. Чтобы разобраться в полученных результатах, полезно рассмотреть термодинамику простой реакции обмена, где липид одного типа вытесняется с места связывания на белке другим липидом:
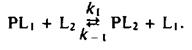
Константа равновесия этой реакции равна
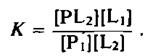
Она является относительной константой связывания липидов Li и L2 с данным участком белковой молекулы, причем
![]()
Если липиды двух типов присутствуют в мембране в одинаковой концентрации и сродство их к белку одинаково, то К = 1. Рассмотрим гипотетический случай, когда L2 является минорным липидным компонентом и составляет только 5°/о от общего количества липидов. Тогда
![]()
Предположим, что L2 предпочтительно связывается с неким участком белковой молекулы, так что в равновесии 90% участков занято L.2. Тогда
![]()
Следовательно, К = 171, что соответствует величине ДС° = = -3,1 ккал/моль. Столь небольшого различия в свободной энергии связывания оказывается достаточно для существенного смещения распределения липидов, связанных с белком, от равновесного состояния. Это обусловлено тем, что соотношение эффективных концентраций конкурирующих липидов в мембране относительно мало: не превышает 100, а в большинстве случаев гораздо меньше. Другими словами, даже минорные липиды присутствуют в мембране в концентрации, составляющей не менее 1% от концентрации основных липидов, с которыми они конкурируют за места связывания на белках.
Для оценки относительного сродства липидов к специфическим белкам используют два подхода. Они основаны на применении липидных аналогов, встроенных в фосфолипидные везикулы, которые содержат интересующий исследователя белок.
1. Спин-меченные фосфолипиды, соседствующие с мембранными белками, обладают ограниченной подвижностью. Это проявляется в уширении спектра ЭПР. У тех молекул ЭПР-зонда, которые соседствуют с белком, возможность движения с характерной частотой > 108 с" 1 существенно ограничена. Спектр ЭПР в этом случае может быть представлен в виде суммы двух компонент: компоненты с узкими спектральными линиями, соответствующей основной липидной фазе, и компоненты, отвечающей липидам с ограниченной подвижностью. Для того чтобы эта последняя была в спектре преобладающей, отношение белок/липид должно быть достаточно высоким. Осложняет картину то, что липиды могут попадать в «ловушки» из белковых агрегатов. Такие липиды тоже обладают ограниченной подвижностью, и им соответствует третья компонента в спектре ЭПР. Для выявления попавших в ловушку липидов при высоких концентрациях белка можно использовать липидные спиновые зонды, ковалентно связанные с поверхностью белка.
2. Спин-меченные и бромированные липидные производные способны тушить флуоресценцию триптофана, входящего в состав мембранных белков. Эффективность тушения зависит от расстояния между липидным производным и остатком триптофана, и присутствие таких липидов в слое, непосредственно прилегающем к белку, приведет к тушению флуоресценции белка. Относительную способность этих липидов связываться с белком в присутствии различных конкурирующих липидов можно исследовать путем измерения интенсивности флуоресценции белка.
Анализ экспериментальных данных, но на практике такие измерения трудно осуществить. Как правило, считается, что все места связывания одинаковы. В одном из вариантов исследуют связывание небольшого количества спин-меченного липида данного типа в присутствии большого количества липида другого типа. При этом снимаются спектры образцов с разным соотношением между липидами и белками. В предположении, что имеется N одинаковых мест связывания, выполняется следующее соотношение. Трудности здесь могут возникнуть из-за гетерогенности мест связывания как по сродству к липидам, так и по способности связанных липидов тушить флуоресценцию триптофана. Однако флуоресцентный метод имеет то преимущество, что при высоких значениях отношения липид/белок агрегация белка создает меньшие проблемы, чем при использовании метода ЭПР.
Необходимо помнить, что ни один из методов, используемых для нахождения относительных констант связывания липидов с мембранными белками, не пригоден для определения небольших количеств мест связывания с высоким сродством, поэтому тот факт, что такие места этими методами не обнаружены, вовсе не означает, что они отсутствуют.
Некоторые белки проявляют избирательность при связывании с фосфолипидами, несущими различные полярные головки
Избирательность связывания с липидами была изучена только для небольшого числа белков, в частности для цитохром с-оксидазы митохондрий, Na+/К+ -АТРазы, Са2 + -АТРазы из саркоплазматического ретикулума и родопсина позвоночных у животных. Как правило, избирательность связывания весьма слаба, при этом максимальное значение К близко к 5. Тем не менее, этого достаточно для того, чтобы имелось значительное различие в связывании липидов с разными белками, зависящее от концентрации липидов разных типов, присутствующих в мембране.
1.Одна молекула родопсина связывается примерно с 24 липидными молекулами, отдавая лишь очень небольшое предпочтение кардиолипину перед фосфатидилхолином.
2.Ыа + /К+-АТРаза из Squalus acanthus связывается примерно с 60 фосфолипидными молекулами, отдавая предпочтение отрицательно заряженным липидам по сравнению с фосфатидилхолином: для кардиолипина К = 3,8, для фосфа-тидилсерина К = 1,7, для фосфатидной кислоты К = 1,5. Фосфатидилэтаноламин и фосфатидилглицерол связываются с этим белком с одинаковым сродством. Предпочтение кислым липидам не может быть обусловлено чисто электростатическими эффектами. Какова биохимическая роль такого предпочтения — неизвестно.
3.Цитохром с-оксидаза связывает от 40 до 55 молекул липидов. Этот фермент отдает предпочтение кардиолипину по сравнению с фосфатидилхолином. Как правило, при очистке митохондриальной цитохром с-оксидазы она выделяется вместе с кардиолипином. Высказывалось предположение, что у этого фермента имеется небольшое число мест связывания, обладающих высоким сродством к этому липиду, и что последний необходим для обеспечения его каталитической активности. Для выяснения биологической роли предпочтительного связывания кардиолипина необходимо провести дальнейшие исследования. Кроме того, поскольку кардиолипин является димерным фосфолипидом, возникают трудности при анализе данных по конкурентному связыванию.
4. Са2 + -АТРаза привлекла внимание ученых в связи с тем, что, как предполагается, ее ферментативная активность определяется так называемым «липидным кольцом», или «пограничным липидным слоем», непосредственно примыкающим к белку. Активность Са2 + -АТРазы не зависела от содержания холестерола в реконструированных везикулах, а чистый холестерол тоже не оказывал никакого влияния на ее активность. Это наводило на мысль, что холестерол в указанном пограничном слое отсутствует. Однако работы по связыванию свидетельствовали об обратном, а дело было в том, что относительная константа связывания холестерола с белком примерно вдвое меньше, чем для фосфатидилхолина. Небольшое количество холестерола присутствует и в природных мембранах, содержащих данный фермент. Относительное сродство Са2 + -АТРазы к фосфо-липидам почти не зависит от природы полярной головки, а также типа ацильных цепей, хотя при реконструкции фермента с разными липидами ферментативная активность существенно варьирует.
Скорость обмена пограничных липидов со свободными липидами
Термин «пограничный липидный слой» был введен в связи с появлением идеи о существовании стационарного слоя липида, связанного с поверхностью белка, например цитохромокси-дазы. Под «стационарностью» понимается отсутствие обмена липидов за время, сравнимое со временем оборота фермента, т. е. ~ 10 ~3 с, поэтому такой фермент вместе с его непосредственным окружением может рассматриваться как долгоживущий комплекс. Во всех изученных к настоящему времени случаях это условие не выполняется, но термин «пограничный липидный слой» остается полезным описательным термином, если речь идет о ближайшем липидном окружении белка.
Данные 2Н-ЯМР четко показали, что пограничные липиды быстро обмениваются с основной массой липидов в бислое. Спектры ЭПР меченых фосфолипидов свидетельствуют о существовании свободных и связанных липидных компонентов, в то же время аналогичные эксперименты с дейтерированными липидными зондами говорят о том, что организация и динамические свойства липидов в присутствии таких белков, как цитохромоксидаза, практически не изменяются. Такое расхождение, по-видимому, объясняется различиями в характерных временах измерения двух методов. Время обмена пограничных липидов составляет 10"6— 10 7 с, поэтому метод 2Н-ЯМР выявляет только один тип липидов, тогда как данные ЭПР свидетельствуют о наличии двух разных липидных популяций.
Если коэффициент латеральной диффузии фосфолипидов в мембранном бислое равен величине, приведенной в табл. 5.2, то среднее время перескока липидной молекулы из одного положения в мембране в соседнее составляет около 10"7 с. Наличие такого быстрого обмена между пограничными и свободными липидами позволяет предположить, что в известных нам случаях липиды, находящиеся по соседству с белком, и свободные липиды практически равноценны. Необходимо подчеркнуть, что это только грубые оценки, полученные для относительно небольшого числа белков, главным образом производных фосфатидилхолина.
5.2 Изменения в липидном бислое, связанные с присутствием интегральных мембранных белков
Результаты большинства исследований липидных бислоев методом 2Н-ЯМР с применением дейтерированных фосфолипидов свидетельствуют о том, что белки оказывают лишь слабое влияние на упорядоченность липидов в бислое и их динамические свойства. Сходные данные были получены методами 'Н-, l9F- и 3|Р-ЯМР. Имеются сообщения о существовании пограничных липидов в природных мембранах. Эти данные получены методом 3,Р-ЯМР и предполагают иммобилизацию полярных головок дол-гоживущими липидно-белковыми комплексами. Однако эти факты нельзя считать окончательно установленными. Так, из данных ЯМР можно сделать следующие общие выводы: 1) скорость обмена между пограничными и свободными липидами весьма велика; 2) в присутствии белков упорядоченность связанных липидов почти не изменяется; 3) скорость переориентации ацильных цепей в присутствии белков слабо уменьшается в частотном диапазоне 109с~'; 4) соседство с трансмембранными белками не сказывается существенным образом на ориентации и динамических свойствах полярных головок. По данным ЯМР, организация и динамика липидов при контактировании с белками очень слабо, но изменяются. Это связывают с некой «жесткостью» поверхности белка, контактирующего с липидом, но сами эффекты весьма незначительны.
Влияние мембранных белков на липидные бислои исследовали и многими другими методами. Рассмотрим вкратце некоторые из полученных результатов.
1. Подвижность спин-меченных липидов ограничивается при контактировании их с белками или при захвате липидно-белковыми агрегатами. В большинстве ЯМР-исследований по релаксации сходные эффекты в том же частотном диапазоне не обнаруживаются. Возможно, эти эффекты обусловлены присутствием объемной нитроксидной спиновой метки. Однако, по крайней мере, в одном случае и данные ЯМР, и данные ЭПР свидетельствовали об уменьшении подвижности ацильных цепей в этом временном диапазоне при липидно-белковых взаимодействиях. Единого мнения о том, влияет ли белок на подвижность спиновых меток за пределами пограничного липидного слоя, не выработано. О существовании некоего липидного слоя за пределами пограничного слоя свидетельствовали спектроскопические данные; эти данные объяснялись в предположении, что имеется единый липидный слой, подверженный воздействию белков. В любом случае важно помнить, что методом ЯМР значительные ограничения подвижности обычно не выявляются.
2.Деполяризация флуоресценции мембранных зондов, например ДФГ, зависит от присутствия интегральных мембранных Щелков. По мере встраивания белков в бислой микровязкость последнего возрастет, о чем свидетельствуют поляризационные данные в стационарных условиях. Для согласования экспериментальных данных с представлением о том, что ограничение подвижности испытывают только зонды, соседствующие с белком, использовалось простое моделирование. Однако детальный анализ, Включавший измерения анизотропии образцов во времени, в одном случае показал, что а) влияние белка проявляется в основном в увеличении параметра упорядоченности, т. е. в ограничении диапазона Движения, и б) данный эффект распространяется за пределы пограничного слоя. Однако эффекты, наблюдаемые при работе с этими зондами, не обязательно будут характерны для фосфолипидов.
3.Для определения конфигурации ацильной цепи использовался Метод инфракрасной спектроскопии с фурье-преобразованием. Полученные результаты согласовывались с данными 2Н-ЯМР и свидетельствовали о незначительном изменении упорядоченности ацильной цепи в присутствии белка. Природа наблюдаемых эффектов зависит от свойств ацильной цепи, поэтому построение какой-то общей модели затруднено {34]. Использование дейтерированных препаратов позволяет избирательно изучать различные липиды; так, было выявлено преимущественное взаимодействие между гликофорином и фосфатидилсерином.
4. Для изучения температурного фазового перехода в липидах и влияния мембранных белков на этот переход использовалась дифференциальная сканирующая калориметрия. Вообще говоря, в присутствии интегральных мембранных белков происходят: а) весьма незначительное уменьшение температуры фазового перехода; б) увеличение ширины интервала перехода и в) уменьшение АН перехода. Если в присутствии белка наблюдается только один фазовый переход, то его относят к липидам, которые не подвергаются влиянию белков. Белок как бы изолирует связанные с ними липиды и предотвращает их участие в фазовом переходе. Это приводит к уменьшению молярной энтальпии перехода. Обычно АН0 уменьшается с увеличением содержания белка линейно, что позволяет легко найти число липидных молекул, связанных с одной молекулой белка. Это число варьирует от примерно 20 для бактериородопсина до 685 для белка полосы 3. Сюда входят не только молекулы пограничных липидов, но и липиды, захваченные белковыми агрегатами, находящиеся в обогащенных белком доменах, а также, возможно, липиды, претерпевшие изменения из-за взаимодействий с углеводными цепями гликопротеинов. Латеральное разделение фаз и агрегация белков затрудняют использование этого подхода для получения подробной информации о липидно-белковых взаимодействиях. Однако изучение влияния белков на температурные переходы бинарных смесей липидов позволило выявить преимущественные взаимодействия между гликофорином и фосфатидилсерином и между цитохром с-оксидазой и кардиолипином.
В некоторых случаях были зарегистрированы дополнительные индуцированные белком фазовые переходы в липидах. Возможно, эти липиды содержатся в обогащенных белками доменах, а может быть, наблюдаемый эффект связан с влиянием изолированных белковых молекул белка на липиды, не принадлежащие пограничному слою.
Влияние белков на полиморфизм фосфолипидов
Некоторые белки оказывают сильное влияние на полиморфизм фосфолипидов, стабилизируя ламеллярные или гексагональные формы. Этот факт представляет интерес в связи с гипотезой о том, что инвертированные гексагональные цилиндры или липидные частицы играют какую-то роль в перемещении липидов через мембранный бислой и в слиянии мембран. В присутствии гидрофобного полипептида грамицидина А при соотношении липид/ белок, равном 10:1, диолеилфосфатидилхолин переходит из бислойной структуры в гексагональную Нц-фазу. При этом, по-видимому, происходит агрегация грамицидина и последующая дегидратация липидов, стабилизирующая гексагональную фазу. Напротив, гликофорин стабилизирует бислойную конфигурацию диолеилфосфатидилэтаноламина, в то время как обычно этот липид находится в гексагональной Нц-фазе. На полиморфизм кардиолипина влияют положительно заряженные белки, например кардиотоксин, цитохром с. В связывании кардиотоксина участвуют как электростатические, так и гидрофобные силы, как и в случае белков, связывающихся с поверхностью мембраны.
Возможная роль упругих деформаций бислоя, обусловленных белками
Искажения в структуре бислоя, о которых шла речь в предыдущем разделе, в большинстве своем возникают при точном соответствии ацильных цепей липидов форме белковой молекулы, при этом налагаются некоторые ограничения на определенные быстрые движения липидных молекул, соседствующих с белком. Такие искажения распространяются на очень небольшие расстояния, лишь немного выходя за рамки пограничного слоя. Интересно было бы рассмотреть более серьезные возмущения, при которых белки, встраиваясь в бислой, производят более существенные изменения в липидном бислое. Некоторые возмущения такого рода представлены на рис. 5.7. В принципе деформации, индуцируемые в бислое, могут распространяться на значительные расстояния, так реорганизуя липиды и/или белковые компоненты, что система переходит в наиболее стабильное состояние. Было разработано несколько теоретических подходов к исследованию этого вопроса, но, к сожалению, экспериментальные данные весьма немногочисленны.
1. Белок в форме клина или белок, проникающий только в один монослой, изменяет наклон ацильных цепей липидных молекул в одном или обоих слоях мембраны. Это изменение может распространяться на большие расстояния от белка и влиять на взаимодействия липидов с другими мембранными белками. Возникающие при этом напряжения в бислое могут сниматься благодаря реорганизации липидов. Например, липиды с относительно небольшими полярными головками могут группироваться вокруг белковой молекулы. Одним из преимуществ многокомпетентности мембраны может быть оптимизация упаковки липидов
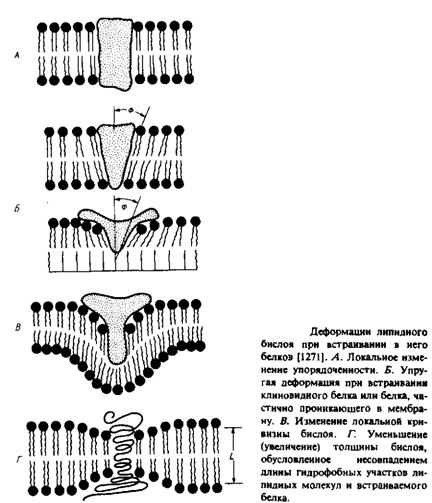
вокруг отдельных мембранных белков, уменьшающая возможные деформации на границе белок—липид.
2.Другой тип деформаций, возникающих при встраивании белка в мембрану, — это латеральные искривления. Примером такого белка может служить «непрочно» связывающаяся форма цитохрома.
3.Еще одной причиной деформаций бислоя может служить несоответствие между размером данного гидрофобного участка бислоя и толщиной мембраны. Чтобы избежать экспонирования гидрофобных областей в воду, белок или липиды могут частично изменить свою конформацию. Если белок не деформируется, то может произойти следующее: а) или ацильные цепи, или белковая молекула наклоняются относительно нормали к бислою на угол, зависящий от толщины мембраны. Это предположение было в одном случае подвергнуто проверке и не нашло подтверждения; б) ацильные цепи липидов деформируются; в) в гетерогенной смеси липидов последние реорганизуются таким образом, что молекулы «неправильной» длины оказываются сгруппированными вокруг белка.
В принципе подобные упругие деформации могут индуцировать специфические взаимодействия липидов с определенными белками для уменьшения искажений в структуре бислоя путем подгонки формы и размера этих молекул, а не за счет специфических химических взаимодействий. Однако эксперментальные данные на этот счет отсутствуют. Кроме того, простирающиеся на большие расстояния деформации могут влиять на белок-белковые ассоциаты. Экспериментальные подтверждения этому были получены в результате наблюдения с помощью электронной микроскопии за распределением бактериородопсина и родопсина в реконструированных фосфолипидных везикулах при разной толщине мембраны. Адаптированный к темноте родопсин действительно агрегировал в результате изменения наклона ацильных цепей липидов при внедрении фермента в бислой, слишком толстый для идеальной упаковки вокруг белка.
5.3 Динамические свойства остова мембранных белков и их боковых цепей
Проводя ЯМР-исследования твердых образцов, можно получить детальную информацию о динамических свойствах отдельных аминокислотных остатков мембранных белков. Однако при этом необходимы большие количества препарата равномерно меченного белка. Наиболее информативным этот метод является в случае небольших белков, когда можно проводить спектроскопические измерения. Возможности этих методов иллюстрируют работы по исследованию белков оболочки нитевидных бактериофагов. При вирусной инфекции эти белки встраиваются в плазматическую мембрану Е. соН с помощью единственной трансмембранной спирали, а во время сборки фага липиды и белки клетки-хозяина исключаются из его оболочки. Малый размер белков оболочки, возможность получения их в больших количествах и легкость выделения создают значительные преимущества при их изучении методом ЯМР, а также другими методами.
Для исследования динамических свойств аминокислотных остатков белка оболочки фага fd в реконструированных фосфолипидных бислоях использовались методы 2Н- и 15N-flMP. Результаты показали, что полипептидный остов на участке от 5-го до 43-го остатков включительно относительно жесткий, при этом данный сегмент превосходит по длине участок, находящийся внутри липидного бислоя. Несколько остатков на концах полипептида свободны и могут совершать движения с большой амплитудой. Большинство боковых цепей в состоянии совершать такие движения, даже когда соответствующие остатки находятся внутри бислоя.
Работы, в которых использовались упомянутые методы, слишком немногочисленны для того, чтобы можно было составить полное представление о характере влияния липидно-белковых взаимодействий на внутреннюю динамику мембранных белков.
5.4 Связывание периферических мембранных белков с липидным бислоем
При изучении липидно-белковых взаимодействий основное внимание уделялось трансмембранным белкам, однако в последнее время проявляется все больший интерес к связыванию с бислоем периферических мембранных белков. Многие такие белки связываются с мембраной главным образом через взаимодействие с интегральными белками. Но существует большая группа разнообразных белков, которые связываются непосредственно с поверхностью липидного бислоя. Некоторые из этих белков, например основный белок миелиновой оболочки, спектрин и матриксный белок вируса везикулярного стоматита, играют в основном структурную роль. Множество растворимых белков связываются с поверхностью мембраны на непродолжительное время или при специфических условиях. В некоторых случаях связывание белка является необходимым условием проявления его ферментативной активности; такими белками являются, например, протеинкиназа С, факторы свертывания крови, пируватоксидаза. Еще одним примером белков, связывающихся с поверхностью бислоя, служат амфифильные пептидные гормоны и, возможно, сигнальные последовательности, которые ответственны за перемещение секретируемых или мембранных белков в нужное место.
По-видимому, существует два основных, не исключающих друг друга типа связывания белков с липидами: 1) связывание осуществляется при участии амфифильной структурной единицы, обычно а-спирали. Эта вторичная структура может индуцироваться и стабилизироваться при взаимодействии с липидами; 2) связывание имеет в основном электростатическую природу и осуществляется при участии положительно заряженного участка белковой молекулы и кислых фосфолипидов. При этом значительную роль могут играть гидрофобные взаимодействия, зависящие от того, насколько глубоко белок проникает в бислой. Во многих случаях для связывания с кислыми фосфолипидами необходим Са2 +, но истинная роль этого двухвалентного катиона точно не определена.
Взаимодействие периферических мембранных белков с фосфолипидами изучали многими методами. Так, за связыванием белков с везикулами можно следить с помощью светорассеяния или путем измерения флуоресценции белков, при этом можно определить константы диссоциации. Возмущения в бислое, вызванные связыванием с ним белков, можно выявить по изменению проницаемости везикул или параметров температурного фазового перехода липидов, хотя анализировать эти результаты на молекулярном уровне довольно трудно. Весьма полезным оказалось также изучение монослоев, при этом степень проникновения белка в монослой можно оценить по изменению площади поверхности монослоя после внедрения белка.
Для получения детальной информации на молекулярном уровне одним из наиболее ценных методов оказался ЯМР. С помощью ЯМР были детально проанализированы последствия взаимодействия липидного бислоя с основным белком миелиновой оболочки и цитохромом с. Оба этих белка взаимодействуют с кислыми липидами главным образом электростатически, хотя физиологическая роль такого взаимодействия цитохрома с с липидами неясна. В отличие от трансмембранных белков два указанных периферических белка значительно различаются по взаимодействию с фосфолипидами. Так, при изучении везикул, содержащих димиристоил-фосфатидилглицерол и фосфатидилхолин, обнаружилось, что основный белок миелиновой оболочки специфически взаимодействует с первым из этих липидов. Исследования методом инфракрасной спектроскопии с фурье-преобразованием показывают, что при связывании с фосфатидилглицеролом белок приобретает высокоупорядоченную вторичную структуру; в основном он образует /3-слой, который в отсутствие этого липида не наблюдается. По данным 2Н-ЯМР белково-липидные взаимодействия приводят к существенному изменению упаковки полярных головок кислых фосфолипидов. Сходные работы, выполненные на цитохроме с с использованием метода 2Н-ЯМР, показали, что при связывании с этим белком происходят лишь небольшие изменения в упаковке головок фосфатидилсерина; в этих опытах использовались везикулы, содержащие также фосфатидилхолин. В обеих системах не наблюдалось никакого латерального разделения фаз и происходил быстрый обмен между свободными и связанными с белками липидами. Однако с другими кислыми фосфолипидами цитохром с взаимодействует по-разному. Например, в везикулах, содержащих кардиолипин и фосфатидилхолин, он вызывает латеральное разделение фаз, а в везикулах, содержащих кардиолипин и фосфатидилэтаноламин, стабилизирует иебислойные структуры.
Итак, по-видимому, типы взаимодействия между периферическими мембранными белкам и фосфолипидным бислоем весьма разнообразны.
Резюме
Чтобы до конца установить функции биологических мембран, необходимо изучить динамические свойства их компонентов. Для измерения скорости вращения липидов и белков внутри мембраны и скорости латеральной диффузии этих компонентов в плоскости мембраны были разработаны специальные спектроскопические методы. Они основаны на использовании спиновых или флуоресцентных зондов, которые встраиваются в мембрану или связываются с конкретными белками. Как правило, мембранные липиды могут свободно диффундировать в плоскости мембраны со скоростью, сравнимой со скоростью их диффузии в модельных мембранах. Напротив, латеральное движение интегральных белков в биологических мембранах часто ограничено. Это может быть связано с их ассоциацией с другими мембранными белками или с элементами цитоскелета либо внеклеточного матрикса. Многие белки способны свободно вращаться в плоскости мембраны, но это вращение также может быть затруднено из-за образования белковых агрегатов.
В любой момент времени с белком в биологической мембране соседствует значительная доля липидов. Слой липидов, непосредственно прилегающих к белку, называется пограничным. Эти липиды очень быстро обмениваются с основной массой липидов бислоя, и обычно вероятность нахождения тех или иных липидов по соседству с белком или в основной липидной фракции почти одинакова. Правда, для некоторых белков характерна определенная избирательность в связывании с липидами, но для всех известных случаев различие в связывании, измеряемое сродством липидов к пограничному слою, не превышает пяти. И даже такая слабая избирательность может приводить к тому, что липидный состав пограничного слоя будет отличаться от состава основной липидной фазы.