
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
Психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Курсовая работа: Происхождение жизни на Земле
Курсовая работа: Происхождение жизни на Земле
Содержание
Введение………………………………………………………..........…………….2
1. Современные представления о происхождении жизни на Земле.......………4
2. Основные законы биологической эволюции…………………………..........18
3. Микро и макроэволюция………………………………………………..........22
Заключение……………………………………………………………….............32
Список используемой литературы………………………………………...........33
Введение
С момента возникновения жизни природа находится в непрерывном развитии. Процесс эволюции продолжается уже сотни миллионов лет, и его результатом является то обилие форм живого, которое во многом до конца еще не описано и не классифицировано. Эволюция протекает на всех уровнях организации живой материи и на каждом уровне характеризуется новообразованием структур и появлением новых функций. Объединение структур и функций одного уровня сопровождается переходом живых систем на более высокий эволюционный уровень.
Существует много определений понятия "жизнь" - столь же сложного, многогранного и неоднозначного, как понятия "Вселенная", "материя" и "разум", предельно широкого, отражающего самые общие черты действительности - категории, определяемой через описание основных характеристик и свойств.
Определение из энциклопедического словаря весьма уязвимо: "Живыми называются системы, которые способны самостоятельно поддерживать и увеличивать свою очень высокую степень упорядоченности в среде с меньшей степенью упорядоченности" - по нему живыми являются все самоорганизующиеся и саморегулирующиеся системы - звезды, галактики и сама Метагалактика, коацерватные капли и многие сложные органические соединения, самопроектирующиеся и самосборные кибернетические устройства и т.д.. Предложение академика С.Ф. Лихачева рассматривать жизнь как некоторое неопределимое свойство Вселенной ничего не дает в плане практического использования понятия.
Не потеряло своей актуальности уточненное в ХХ веке определение Ф. Энгельса: "Жизнь - это способ существования белковых тел и нуклеиновых кислот" – поскольку вне живых организмов белки в природе не встречаются.
Наилучшим следует признать определение академика Н.В. Волькенштейна: "Живые тела, существующие на Земле, представляют собой открытые, самоорганизующиеся и самовоспроизводящие системы, построенные из биополимеров - белков и нуклеиновых кислот".
Проблемы происхождения и эволюции жизни на Земле принадлежали и принадлежат к числу величайших проблем естествознания. Эти проблемы привлекали к себе внимание человеческого ума с самых незапамятных времен. Они являлись предметом интереса всех философских и религиозных систем. Однако в разные эпохи и на разных ступенях развития человеческой культуры проблемы происхождения и эволюции жизни решались по-разному. Сейчас считается общепризнанным, что возникновение жизни на Земле представляло собой закономерный процесс, вполне поддающийся научному исследованию. Однако вопрос о происхождении жизни труден в исследовании, поскольку когда наука подходит к проблемам развития как сотворения совершенно нового, она оказывается у предела собственных возможностей, так как ученые сейчас не в состоянии воспроизвести процесс возникновения жизни с такой же точностью, как это было несколько миллиардов лет назад. Даже тщательно поставленный опыт будет только модельным экспериментом, лишенным ряда факторов, сопровождавших появление живого на Земле.
Таким образом, вопрос о происхождении жизни интересен не только сам по себе, но и тесной связью с вопросами эволюции форм жизни.
Актуальность выбранной темы не вызывает сомнений, поэтому в данной работе рассматриваются современные теории возникновения жизни и процесса эволюции ее форм.
При выполнении работы использовались учебники и учебные пособия по концепциям современного естествознания, философии, астрономии, экологии, экологическому праву и природопользованию, а также монографии и научные статьи в периодических изданиях.
1. Современные представления о происхождении жизни на Земле
В настоящее время ученые выделяют пять научных концепций возникновения жизни:
1. Возникновение живого из неживого, подчиняясь определенным физическим и химическим закономерностям - абиотическая концепция;
2. Гипотеза «голобиоза» - концепция протобионта либо биода, некоего доклеточного предка, начальных «жизнеспособных» структур;
3. Гипотеза «генобиоза», т.е. поиска генома как реликтового предка всех живых клеточных структур, считая, что конкретно РНК сыграла первостепенную роль в ёе зарождении жизни;
4. Концепция стационарного состояния жизни - жизнь была постоянно, начала жизни не существует;
5. Внеземное происхождение жизни – жизнь была занесена на Землю из Космоса (концепция панспермии).
В развитии учений о происхождении жизни существенное место занимает теория, утверждающая, что все живое происходит лишь от живого - теория биогенеза. Эту теорию в середине XIX века противопоставляли ненаучным представлениям о самозарождении организмов. Но как теория происхождения жизни биогенез несостоятелен, поскольку принципиально противопоставляет живое неживому, утверждает отвергнутую наукой идею вечности жизни [7. С. 129].
Абиотическая концепция. Абиогенез - мысль о происхождении живого из неживого - начальная гипотеза современной теории происхождения жизни. В 1924 г. биохимик А.И.Опарин высказал предположение, что при массивных электрических разрядах в земной атмосфере, которая 4-4,5 млрд. лет назад состояла из аммиака, метана, углекислого газа и паров воды, могли появиться простые органические соединения, нужные для возникновения жизни. Предсказание академика Опарина оправдалось. В 1955 г. американский исследователь С.Миллер, пропуская электрические заряды через смесь газов и паров, получил простые жирные кислоты, мочевину, уксусную и муравьиную кислоты и несколько аминокислот. Таковым образом в середине XX века был экспериментально осуществлен абиогенный синтез белковоподобных и других органических веществ в условиях, воспроизводящих условия первобытной Земли.
Гипотеза Опарина о возникновении жизни на Земле опирается на представление о постепенном усложнении химической структуры и морфологического вида предшественников жизни (пробионтов) на пути к живым организмам. На стыке моря, суши и воздуха создавались благоприятные условия для образования сложных органических соединений. В концентрированных растворах белков, нуклеиновых кислот могут образовываться сгустки подобно акварастворам желатина. А.И.Опарин назвал эти сгустки коацерватными каплями либо коацерватами.
Коацерваты - это обособленные в растворе органические многомолекулярные структуры. Это еще не живые существа. Их возникновение рассматривают как стадию развития преджизни.
Более принципиальным этапом в происхождении жизни было возникновение механизма воспроизведения себе подобных и наследования параметров прошлых поколений. Это стало возможным благодаря образованию сложных комплексов нуклеиновых кислот и белков. Нуклеиновые кислоты, способные к самовоспроизведению, стали контролировать синтез белков, определяя в них порядок аминокислот, а белки-ферменты осуществляли процесс сотворения новейших копий нуклеиновых кислот. Так появилось основное свойство, характерное для жизни - способность к воспроизведению подобных себе молекул.
Сильная сторона абиогенетической гипотезы - её эволюционный характер (жизнь – закономерный этап эволюции материи), возможность экспериментальной проверки главных положений гипотезы (на коацерватных каплях можно сымитировать доклеточные фазы зарождения жизни). Слабая сторона гипотезы Опарина допускала воспроизводство протоживых структур в отсутствии молекулярных структур генетического кода. Гипотеза Опарина предъявляет особые требования к экспериментальному воспроизведению коацерватных структур: «первичный бульон» с химически сложной структурой, элементы биогенного происхождения (ферменты и коферменты).
Абиогенная гипотеза встретила решительный отпор ученых – приверженцев идеи вечности и безначальности биологической жизни. Российский ученый биохимик С.П. Костычев в брошюре «О появление жизни на Земле» замечает, что простые организмы посложнее всех фабрик и заводов, и случайное возникновение жизни маловероятно, жизнь никогда «не создается из мертвой материи» [7. С. 132].
В отношении самозарождения организмов нужно отметить, что Французская Академия наук еще в 1859 г. назначила специальную премию за попытку осветить по-новому вопрос о самопроизвольном зарождении жизни. Эту премию в 1862 г. получил известный французский ученый, основатель современной микробиологии Л.Пастер. Своими опытами он доказал невозможность самозарождения микроорганизмов.
Следует отметить, что в настоящее время жизнь на Земле не может появиться абиогенным методом. Еще Дарвин в 1871 г. писал: "Но если бы сейчас в каком-либо теплом водоеме, содержащем все нужные соли аммония и фосфора и доступном действию света, тепла, электроэнергии и т.п., химически образовался белок, способный к дальнейшим все более сложным превращениям, то это вещество немедленно было бы разрушено и поглощено, что было нереально в период возникновения живых существ".
С гипотезой А.И.Опарина в настоящее время трудно согласиться. Наличие аналогов обмена веществ и «естественного отбора» у коацерватов еще не есть доказательство того, что они могли привести к образованию первых примитивных организмов. А как произошел качественный скачок от неживого к живому гипотеза Опарина не объясняет.
Теория панспермии. Идея панспермии была выдвинута в 1907 году С. Аррениусом и поддерживалась такими крупными учеными, как Э. Хойл, Ч. Викрамсинх, У. Крик, С. Ортель, К. Саган. В 1865 г. немецкий врач Г.Рихтер выдвинул гипотезу космозоев (космических зачатков), в согласовании с которой жизнь является вечной и зачатки, населяющие мировое пространство, могут переноситься с одной планеты на другую. Сходную гипотезу в 1907 г. выдвинул шведский естествоиспытатель С.Аррениус, предположив, что во Вселенной постоянно есть зародыши жизни. Он описывал, как с населенных другими существами планет уходят в мировое пространство частички вещества, пылинки и живые споры микроорганизмов. Они сохраняют свою жизнеспособность, летая в пространстве Вселенной за счет светового давления. Попадая на планету с подходящими условиями для жизни, они начинают новенькую жизнь на данной планете. Эту гипотезу поддерживали многие, в т.ч. российские ученые С. П. Костычев, Л.С. Берг.
Данная гипотеза не предполагает никакого механизма для объяснения первичного возникновения жизни и переносит дилемму в другое место Вселенной. Либих считал, что «атмосферы небесных тел, а также вращающихся космических туманностей можно считать как вековечные хранилища оживленной формы, как вечные плантации органических зародышей», откуда жизнь рассеивается в виде этих эмбрионов во Вселенной. Схожим образом мыслили немецкие ученые Кельвин, Гермгольц.
Для обоснования панспермии традиционно употребляют наскальные картинки с изображением предметов, похожих на ракеты либо космонавтов, либо появления НЛО. Полеты космических аппаратов разрушили веру в существование разумной жизни на планетах солнечной системы, которая возникла после открытия Скипарелли каналов на марсе в 1877 г. [7. С. 135]
Ловелл насчитал на Марсе 700 каналов. Сеть каналов обхватывала все материки. В 1924 г. каналы были сфотографированы, и большая часть ученых узрели в них подтверждение существования разумной жизни. Фотоснимки 500 каналов зафиксировали и сезонные конфигурации цвета, которые подтвердили идеи русского астронома Г.А. Тихова о растительности на Марсе, поскольку озера и каналы имели зеленоватый цвет. Ценная информация о физических условиях на Марсе была получена русским космическим аппаратом «Марс» и американскими посадочными станциями «Викинг-1» и «Викинг-2». Так, полярные шапки, испытывающие сезонные конфигурации, оказались состоящими из аквапара с примесью минеральной пыли и из жесткой двуокиси углерода (сухого льда). Но пока следов жизни на Марсе не найдено. Исследование поверхности с борта искусственных спутников позволило предположить, что каналы и реки Марса могли появиться в итоге растапливания под поверхностного водяного льда в зонах повышенной активности либо внутреннего тепла планеты, либо при периодических конфигурациях климата.
В конце 60-х годов вновь возник интерес к гипотезам панспермии. При исследовании вещества метеоритов и комет были обнаружены многие «предшественники живого» - органические соединения, синильная кислота, вода, формальдегид, цианогены. Формальдегид, в частности, найден в 60% случаев в 22 исследованных областях, облака с его концентрацией приблизительно 1000 молекул в кубическом сантиметре заполняют обширные пространства. В 1975 г. предшественники аминокислот найдены в лунном грунте и метеоритах [2. С. 76].
Некоторые ученые считают, что жизнь возникла не на Земле, а была занесена на нее из космического пространства в виде спор микроорганизмов размерами 0,2-0,6 мкм. В составе планетарных и диффузных газопылевых туманностей и глобул обнаружены сложные органические соединения. При прохождении Солнечной системы в спиральных рукавах на поверхность Земли может выпадать до 1014 органических молекул на м2.
В пользу вышеупомянутой гипотезы свидетельствуют: универсальность генетического кода всех земных организмов и важность роли молибдена в живой клетке, более редкого на Земле, чем хром и никель, которые могли бы выполнять те же функции. Устойчивые к ультрафиолетовому облучению и космической радиации споры действительно могли бы путешествовать между звезд, но для того, чтобы за миллиард лет Земля получила 1 спору, все остальные звезды Галактики должны иметь планеты, выбрасывающие в тот же срок в космос по 1 т. спор.
В качестве места возникновения и развития жизни предполагались кометы и каменные метеориты - углистые хондриты, содержащие сложные органические вещества, в том числе аминокислоты (в составе метеорита Мерчисон было обнаружено 18 разновидностей аминокислот), не встречающиеся на Земле. Многочисленные данные свидетельствуют об изобилии органических соединений на поверхности Земли в эпоху ее формирования [2. С. 79].
Бактерии способны размножаться в экстремальных условиях температур от – 25њ С до 300њ С при давлении до 1,3× 108 Па. Они сохраняют жизнеспособность в виде спор при температурах от – 240њ С до 600њ С и давлении от 10-4-10-6 Па до 2× 109 Па, облучению ультрафиолетовым излучением интенсивностью до 5× 104 эрг/мм2 и жесткой радиации мощностью до 104 Гр. Колонии бактерий (E. coli) на борту АМС "Сервейер" смогли выжить в течение 1 года на поверхности Луны, и свыше 5 лет находившихся на открытой панели ИСЗ. В ископаемых льдах Арктики и Антарктиды обнаружены споры микроорганизмов (до 107 клеток на грамм), находящихся в состоянии анабиоза от 20-40 тысяч лет до 8 миллионов лет! Некоторые ученые полагают, что у многих обнаруженных популяций микроорганизмов в условиях вечной мерзлоты метаболизм крайне замедляется, но не останавливается. Такая жизнеспособность обусловлена неразрывностью связи популяций организмов со средой обитания.
Отдельные группы земных микроорганизмов, примитивных грибов, дрожжей и водорослей могут не только выжить, но и размножаться в условиях, существующих в криосфере и на поверхности Марса или в океанах Европы. И все же, несмотря на космическую распространенность органических веществ, гипотеза панспермии до сих пор не получила материального подтверждения, хотя массовая печать неоднократно в форме сенсаций сообщала об обнаружении в метеоритах окаменелостей микроорганизмов. Главным недостатком этой гипотезы является то, что перенос места возникновения земной жизни с поверхности Земли в глубины Вселенной не решает вопроса о происхождении жизни из неживой материи.
Концепция стационарного состояния жизни. По мнению В.И. Вернадского, необходимо говорить об извечности жизни и проявлений её организмов, как мы говорим об извечности материального субстрата небесных тел, их тепловых, электрических, магнитных параметров и их проявлений. Далек от научных исканий вопрос о начале жизни, как и вопрос о начале материи, теплоты, электроэнергии, магнетизма, движения. Все живое вышло от живого (принцип Реди). Примитивные одноклеточные организмы могли появиться лишь в биосфере Земли, а шире, в биосфере Вселенной. По мнению Вернадского, естественные науки построены на предположении, что жизнь с её особыми свойствами не играет никакой роли в жизни Вселенной. Но биосферу необходимо брать как целое, как единый живой космический организм (тогда и отпадает вопрос о начале живого, о скачке от неживого к живому) [8. С. 12].
Гипотеза «голобиоза» касается прообраза доклеточного предка и его способностей. Есть разные формы доклеточного предка – «биоид», «биомонада», «микросфера». Согласно биохимику П. Деккера, структурную базу «биоида» составляют «жизнеподобные» неравновесные диссипативные (от лат. «dissipate») структуры, т.е. открытые микросистемы с ферментативным аппаратом, катализирующим метаболизм биоида. Эта гипотеза трактует активность доклеточного предка в обменно-метаболическом духе. В рамках гипотезы «голобиоза» конструировали биохимики С.Фокс и К.Дозе свои биополимеры, способные к метаболизму – комплексному белковому синтезу. Основной недостаток данной гипотезы – отсутствие генетической системы при таком синтезе. Отсюда - следует отдать предпочтение «молекулярному прародителю» всякого живого, а не первичной протоклеточной структуре.
Гипотеза «генобиоза». Американский ученый Холдейн считал, что первичной была не структура, способная к обмену веществ с окружающей средой, а макромолекулярная система, подобная гену и способная к репродукции, а потому и названным им «голым геном». Общее признание гипотеза «генобиоза» получила после открытия РНК и ДНК и их феноменальных параметров. В начале 80-х гг. 20 века была установлена способность РНК к саморепродуцированию в отсутствии белковых ферментов. Второй момент – открытие у РНК автокаталитических функций. Объединение двух функций – каталитической и информационно-генетической – привело к тому, что макромолекулярная система стала способной к саморепродукции. Т.е. старая РНК кооперируя в себе черты фенотипа и генотипа, могла быть подвержена как генетическим преобразованиям, так и естественному отбору, т. е. она эволюционировала.
Итак, РНК сыграла первостепенную роль в зарождении жизни. Но мы же знаем, что современный геном биосферы составляет ДНК, а не РНК. Но как это объяснить? Ревертаза и матричный синтез с ДНК на РНК явились компонентами одного для всего живого доклеточного предка. Но эволюция последнего шла в сторону современной ДНК и утраты им самостоятельных каталитических функций. Таким образом, РНК является той первичной информационной молекулой, которая стояла у истоков жизни.
Можно допустить, что обе нуклеиновые кислоты произошли от одной более примитивной молекулы. Усложняясь и специализируясь в процессе эволюции, эта «прануклеиновая» кислота превратилась в функционально различные типы молекул ДНК и РНК. Возможно, что этой первичной нуклеиновой кислотой могла быть молекула, близкая к более простой РНК. Подобно РНК вируса табачной мозаики она обладала способностью к передаче наследственной информации и к синтезу белка. Возможно также, что вирусы содержащие только одну РНК (филогенетически более раннее образование), следует рассматривать как современные модификации древней, примитивной формы жизни [8. С. 15].
Все это может пролить некоторый свет на пути возникновения и развития живых существ от более простых форм управления и примитивной формы жизни к более сложным формам. Если небелковая («неживая») молекула РНК в подходящей среде образует живые системы, то не на этом ли пути можно обнаружить мостик между неживой и живой природой. Решающее слов в этом вопросе принадлежит различным будущим биохимическим и генетическим исследованиям.
Таким образом, основные гипотезы о происхождении жизни на Земле можно разделить на 3 группы:
1) религиозная гипотеза о "божественном" происхождения жизни;
2) "панспермия" - жизнь возникла в космосе и затем была занесена на Землю;
3) жизнь возникла на Земле в результате естественных процессов.
Рассмотрим более подробно закономерности и химические особенности происхождения жизни на земле.
Возникновение и развитие жизни и разума на Земле подготовлено всем ходом эволюции неживой материи Метагалактики. Закономерность и неизбежность возникновения и развития жизни и разума обусловлена одним из важнейших свойств Метагалактики - "антропным принципом". Существование и развитие объектов Метагалактики обусловлено внутренними динамическими процессами. Все возникающие объекты, от космических пылинок и туманностей, бактерий и людей, звезд, галактик и, по-видимому, всей Метагалактики в целом, являются открытыми неравновесными системами, обменивающимися с окружающей средой веществом и энергией. В ходе эволюции возникает способность к воспроизведению подобных объектов и усвоению ими благоприобретенных признаков и свойств. С увеличением сложности структур упорядоченных систем возрастает их способность к накоплению, запоминанию и хранению информации. Информационная эволюция ускоряет темпы самоорганизации материи и идет в направлении уменьшения возможных наборов элементов, определяющих структуру и функционирование сложных систем. Наборы с наименьшим числом элементов легче восстанавливаются, передаются и тиражируются. Для атомов число возможных комбинаций протонов и нейтронов в атомных ядрах равно числу изотопов (свыше 1500). Для молекул возможное число наборов атомов равно количеству элементов таблицы Менделеева (около 100). Для полимеров в растворах - числу 5 пространственных конфигураций. Для живых организмов - количеству нуклеотидов в ДНК и РНК (4). С увеличением сложности структур возрастает зависимость их существования и развития от физических и химических свойств среды и неизменности внешних условий.
Например, температурные границы существования объектов:
- атомов - от 0,5–1 К до 105 К (температура ионизации);
- молекул - от 2-3 К до 104 К (температура диссоциации);
- твердотельных кластеров (объектов, содержащих минимальное число атомов для проявления всех макроскопических свойств данного вещества) - от 10-15К до 5× 103К;
- микроорганизмов - от 100 К до 700 К;
- человека - от 308 К до 312 К [7. С. 142].
Вышесказанное определяет условия, необходимые и достаточные для проявления и развития жизни, возможное время ее возникновения в Метагалактике и на Земле, основные темпы и направления эволюции живых организмов. В раннюю эпоху существования Метагалактики вплоть до образования галактик, жизнь не могла существовать из-за абсолютно неподходящих внешних условий. Не могла она возникнуть вблизи звезд I поколения, которые, скорее всего, не имеют планетных систем из-за 10-40-кратного дефицита тяжелых химических элементов.
Для образования космических тел с современным химическим составом и соотношением изотопов тяжелых элементов их синтез должен был произойти за 4-6 миллиардов лет до образования Солнечной системы, т.е. не позже 9-11 миллиардов лет назад. Образование тяжелых элементов было особенно интенсивным в период формирования основных галактических структур; в нашем районе Галактики период интенсивного звездообразования закончился к моменту образования Солнечной системы.
Химические условия возникновения и развития жизни определяются составом ее молекулярных основ. Нуклеиновые кислоты ДНК и РНК построены из нуклеотидов, состоящих в свою очередь из сахара, азотистых оснований и фосфата; белки состоят из аминокислот. Все химическое разнообразие жизни на Земле исчерпывается 28 веществами: 20 видов аминокислот, 5 оснований, 2 углеводов и 1 фосфата, элементарный химический состав которых состоит из водорода (37,5%), углерода (29,8%), кислорода (18,3%), азота (11,3%), фосфора (3,1%). Водород - самый распространенный химический элемент, углерод, кислород и азот - самые распространенные из тяжелых химических элементов, способные образовывать огромное число сложных и относительно стабильных молекул (благодаря наличию химически инертных соединений углерода). Кислород - активный окислитель, его соединение с водородом Н2О, вода - широко распространенный универсальный биологический химический растворитель, остающийся в жидком состоянии в широком диапазоне температур, обладающий высокой диэлектрической проницаемостью и теплоемкостью.
Химические условия существования жизни налагают ряд дополнительных требований к физическим характеристикам объектов, на которых они могли бы реализоваться [7. С. 135].
Химический состав объекта должен допускать наличие гидросферы и атмосферы приемлемого состава, состоящей из газов, способствующих возникновению и развитию живых организмов и поддерживающих необходимый энергетический режим (температуры и энергетической освещенности) без резких колебаний вышеупомянутых условий и давления. Например, углекислый газ в атмосфере Земли не только основное сырье для фотосинтеза, но и важнейший инструмент для поддержания температуры атмосферы с оптимальной концентрацией 0,03-0,04%. Масса объекта должна обеспечивать силу тяжести, достаточную для удержания постоянной атмосферы достаточной плотности у поверхности космического тела без перехода атмосферных газов в другие агрегатные состояния.
Орбита космического тела должна лежать в пределах "зоны жизни" данной планетной системы, обеспечивающей достаточную энергетическую освещенность поверхности в приемлемом диапазоне длин волн и иметь малый эксцентриситет во избежание резких колебаний внешних условий на поверхности тела. Объект должен вращаться вокруг своей оси со скоростью, достаточной для установления атмосферной и гидросферной циркуляции и некоторого усреднения физических условий на поверхности.
Всем вышеперечисленным условиям отвечают планетные тела (планетоиды и планеты земной группы) массой от 0,1 до 10 МÅ, входящие в состав планетных систем одиночных, медленновращающихся, обладающих постоянством светимости звезд главной последовательности II и последующих поколений спектральных классов F5-К5. Число планетных тел Галактики с благоприятными условиями для существования жизни определяется формулой:
![]() ,
,
где N* - общее число звезд Галактики (около 2× 109);
fn - доля звезд, имеющих планетные системы (все одиночные медленновращающиеся звезды, от 20 до60 % звезд);
ne - доля звезд, вблизи которых могут быть благоприятные для жизни условия (для звезд классов F5-К5 около 0,01-0,02).
Если в каждой из вышеуказанных планетных систем "обитаема" лишь одна планета, то в настоящее время в Галактике может быть от 40 до 240 миллионов планет, на которых существует жизнь. Даже если по каким-либо причинам вероятность возникновения жизни в сотни и тысячи раз меньше, в Галактике сейчас должны быть сотни тысяч и миллионы населенных планетных тел. Для Галактики это очень маленькая величина. Так, на расстоянии до 5 парсек (16,3 св. года) от Солнца насчитывается 53 звезды, из которых лишь 3 - e Эридана, t Кита и e Индейца - удовлетворяют вышеупомянутым условиям; однако у e Эридана планетная система находится в стадии формирования.
В настоящее время в научных лабораториях подробно исследованы и воспроизведены первые этапы эволюции от "неживой" к "живой" материи:
1. Эволюция малых молекул (CH4, H2O, NH3, CO и т.д.).
2. Образование полимеров.
3. Возникновение каталитических функций.
Ведутся исследования последующего этапа эволюции - самосборки молекул-гиперциклов, возникновению биологических мономеров (аминокислот, азотистых оснований и т.д.) и биополимеров, накоплены определенные сведения по следующему этапу - возникновению мембран и доклеточной организации. К сожалению, весьма далеки от окончательного понимания два важнейших заключительных этапа превращения "неживого" в "живое" - возникновение механизма наследственности и возникновение клетки. Основой жизни в Метагалактике могут являться:
1) Широко распространенные химические элементы IV-VI групп таблицы Менделеева (углерод, кремний, кислород, фтор, азот, фосфор, сера и т.д.), способные образовывать сложные молекулярные цепочки, выполняющие функции органических молекул.
2) Химические соединения (вода H2O, аммиак NH3, смесь воды с аммиаком, сероводород H2S, синильная кислота HCN, фтористый водород HF и т.д.), обладающие свойством быть одновременно кислотой и основанием: они способны стать биологическими растворителями. Водородная связь определяет структуру белков, нуклеиновых кислот и других органических соединений и их возможных аналогов [8. С. 36].
"Аммиачная" жизнь является второй по вероятности распространенности после земной, основанной на соединениях углерода и воде. Аммиак обладает достаточно высокими теплотой плавления, парообразования и теплоемкостью, остается жидким в диапазоне температур от –77,7њ С до –33,4њ С при нормальном давлении; при возрастании давления температура кипения увеличивается (до +132,4њ С при р = 112 атм.). Океаны и моря из жидкого аммиака (или смеси аммиака с водой и гидроксиламином NH2OH) будут так же эффективно смягчать колебания температуры, как гидросфера Земли. Аммиак обладает некоторыми биологическими преимуществами перед водой (большей текучестью, способностью растворять органические соединения и т.д.). "Аммиачная" жизнь может процветать на относительно холодных планетах земной группы и плаентоидах с плотными атмосферами.
В плотных атмосферах планет-гигантов в условиях низких температур (от – 100њ С до - 50њ С) может возникнуть сероводородная жизнь. Жизнь может появиться и на поверхности планетных тел с плотными атмосферами из смеси газов CS2, COS, CH4, N2, Ar, и гидросферами из сернистого ангидрида SO2 (Тзамерзания = – 75,5њ С, Ткипения = - 10,2њ С при р = 1 атм).
Кремний может успешно заменить углерод и быть цепочкообразующим элементом органических систем, молекулы которых основаны на связях Si–O–Si или Si–N–Si. "Кремниевая" жизнь может встретиться на планетах, обладающих очень плотными горячими (Т³ 300њ С) атмосферами, обращающихся на небольшом расстоянии вокруг массивных горячих звезд.
Фтор - довольно редкий химический элемент, но жизнь его основе могла бы существовать на планетах земной группы с атмосферами, содержащими свободный F2 как аналог кислорода, и океанами из фтористого водорода HF (Тзамерзания = – 83,1њ С, Ткипения = + 19,5њ С при р = 1 атм), обращающихся на большом расстоянии вокруг высокотемпературных звезд с максимумом энергетической светимости в УФ-диапазоне.
В следующей части работы рассмотрим основные законы биологической эволюции.
2. Основные законы биологической эволюции
Творцом первой подлинно научной теории эволюции стал великий английский ученый Чарльз Роберт Дарвин (1809-1882). Главным трудом Ч.Дарвина является книга "Происхождение видов путем естественного отбора или сохранение благоприятствуемых пород в борьбе за жизнь" (1859), существенным дополнением к которой служат его книги "Изменение домашних животных и культурных растений" (1869) и "Происхождение человека и половой отбор" (1871).
Эволюционное учение Дарвина состоит из трех разделов, а именно: совокупность доводов в пользу того, что историческое развитие организмов действительно имеет место; положение о движущих силах эволюции; представления о путях эволюционных преобразований. Движущими силами эволюции Ч. Дарвин назвал наследственность, изменчивость и естественный отбор. Он считал, что наследственность и изменчивость позволяют фиксировать изменения и трансформировать их в поколениях.
Изменчивость вызывает разнообразие, а наследственность передает эти изменения потомству. Следовательно, для всех живых существ в результате изменений и скрещиваний характерна наследственная гетерогенность.
Поскольку все организмы очень склонны к интенсивному размножению, то в пределах каждого вида производится потомства больше, чем выживает. Излишнее потомство гибнет в результате борьбы за существование, формы которой очень разнообразны [6. С. 115].
По Ч. Дарвину борьба за существование происходит как между видами, так и внутри видов, причем внутривидовая борьба является более ожесточенной по сравнению с межвидовой, ибо особи обитают в одной местности, нуждаются в одинаковой пище, подвергаются одинаковым опасностям и т.д. В результате борьбы за существование выживают наиболее приспособленные, т.е. те организмы, которые обладают каким-либо признаком, обеспечивающим приспособление.
Следовательно, неизбежным следствием борьбы за существование является естественный отбор. "Сохранение благоприятных индивидуальных различий и изменений и уничтожение вредных я назвал естественным отбором или переживанием наиболее приспособленных (Ч. Дарвин). Ч. Дарвин считал, что естественный отбор является главным механизмом эволюции.
Заслуги Ч. Дарвина перед естествознанием имеют непреходящее значение. Он всесторонне обосновал исторический метод в применении к природе, создал теорию эволюции и изгнал из науки креационизм.
Учение Дарвина касалось эволюции крупных групп организмов, включая все таксоны, в большие отрезки времени, измеряемые геологическими масштабами и на обширных территориях. Следовательно, классический дарвинизм является учением о макроэволюции.
Однако оно оказалось недостаточно разработанным в генетическом плане. Дарвин имел в виду эволюцию особей, но ведь особи живут, как сейчас известно, в популяциях. Поэтому через 8 лет после выхода "Происхождения видов" англичанин Ф. Дженкин поставил такой вопрос: "Если отбор оставляет в живых те особи, которые лишь незначительно отличаются от других, то уже при последующем скрещивании наступает "поглощение" новых признаков, т.к. партнер по скрещиванию, вероятнее всего, не имеет этого нового свойства — произойдет растворение при-J знаков в потомстве".
Дарвин называл этот вопрос кошмаром Дженкина, т.к. никогда не мог дать удовлетворяющий ответ. Дарвинизму длительное время не хватало генетической основы, но генетика очень быстро сама подошла к эволюции. Используя методологию классического генетического анализа, генетики стали анализировать роль отдельных факторов эволюции путем вычленения элементарных единиц и протекающих в них процессов [6. С. 118].
Первый шаг на пути объединения дарвинизма и генетики заключался в законе Дж. Харди-В.Вайнберга, которые в 1908 г. показали, что в популяции при свободном скрещивании, отсутствии мутаций данного гена и отсутствии отбора по данному признаку соотношение генотипов АА, Аа и аа остается постоянным. Это позволило им сформулировать закон, содержание которого сводится к тому, что частоты генов в бесконечно большой панмиксической популяции без давления каких-либо внешних факторов стабилизируются уже после одной смены поколений. Однако таких популяций, как известно, в природе не существует. Поэтому значение закона заключается в том, что накопленные наследственные изменения в генофонде популяций бесследно не исчезают, т.е. частоты генов постоянны.
Исходя из закона Харди-Вайнберга и учитывая влияние отбора и возникновение новых мутаций, в 1926 г. С.С. Четвериков (1880-1959) показал, что в результате спонтанного мутационного процесса во всех популяциях создается генетическая гетерогенность. Другими словами, в популяциях всегда есть мутации, в большинстве случаев в малых количествах. Он показал также, что популяция насыщена мутациями, как губка, и что мутации служат основой (материалом) эволюционного процесса, идущего под действием естественного отбора.
Позднее Н.П. Дубинин и Д.Н. Ромашов (1932) показали, что когда популяции малы, в них происходят явления, получившие название генетико-автоматических процессов (по Н.П. Дубинину, 1931) или дрейфа генов (по С.Райту, 1932). В результате генетико-автоматических процессов изменяются частоты встречаемости генов, устраняются гетерозиготы и появляются гомозиготы. Изолированная популяция становится доминантной гомозиготной или рецессивной гомозиготной. Если дрейфует мутантный летальный ген, это ведет к вымиранию организмов [6. С. 122].
Таким образом, структура популяции зависит не только от появления новых мутаций, но и от простого изменения частоты встречаемости данного гена. Эти и другие генетические исследования связали эволюционную теорию с генетикой.
Современная концепция возникновения жизни на Земле является результатом широкого синтеза естественных наук, многих теорий и гипотез, выдвинутых исследователями различных специальностей.
Огромный интерес представляет решение проблем:
- Почему все белковые соединения в составе живого вещества имеют левую симметрию?
- Однократно или многократно возникала жизнь на Земле, было ли ее возникновение глобальным или локальным явлением?
- Почему жизнь на Земле не возникает из неживого в настоящее время?
- Почему у всех живых существ на Земле белки строятся только из 20 аминокислот из более чем 100 известных науке?
- Может ли возникнуть жизнь в других условиях, на принципиально иной химической основе?
Таким образом Эволюция живых организмов от простейших форм к разумным существам занимает, по-видимому, несколько миллиардов лет - на Земле 3,5 миллиарда лет. Движущей силой эволюции являются мутации и естественный отбор - процессы, носящие статистический характер и обусловленные плавными медленными изменениями условий существования организмов (составом, плотностью и температурой атмосферы и гидросферы, климатом, рельефом, магнитным полем планеты, спектральным составом и уровнем освещенности поверхности и т. д.), причинами которых являются незначительные изменения в действии космических факторов в сочетании с мелкими, периодическими и беспорядочными колебаниями ряда основных характеристик внешней среды, в основе которых, как правило, также лежит действие космических процессов и объектов [1. С. 155].
3. Микро и макроэволюция
Эволюция биомассы на Земле рисуется следующим образом. Разделим существующие организмы на группы:
1) водоросли,
2) морские животные,
3) наземные растения,
4) наземные животные.
Водоросли появились в катархее, и в течение архея и протерозоя их биомасса нарастала до величины порядка сегодняшней, т.е. до 1 млрд. т. Морские животные появились в среднем протерозое, их биомасса очень медленно росла до венда, а в течение кембрия, вероятно, быстро выросла до величины порядка сегодняшней, т.е. до 20-30 млрд. т. Наземные растения появились, возможно, еще в ордовике или даже в кембрии, однако их биомасса стала ощутимой лишь в силуре, а за девон и карбон быстро выросла до колоссальных размеров, порядка нескольких триллионов тонн; в перми она, возможно, даже несколько уменьшилась и приблизилась к современному уровню. Наконец, наземные животные появились только в девоне, их биомасса достигла современной величины, вероятно, еще в палеозое.
Что же касается изменений со временем разнообразия форм организмов, то здесь имеются довольно надежные фактические данные. Они показывают, что в разнообразии форм монотонного нарастания со временем не было, так как при появлении новых форм некоторые из старых отмирали. Так, в кембрии появилось 28 классов животных, в ордовике еще 14 (но часть классов уже отмерла), в силуре еще 3 (но 2 отмерли). Начало верхнего палеозоя ознаменовалось новой вспышкой формообразования - в девоне появилось 9 новых классов (и их число достигло фанерозойского максимума), а карбоне еще 5 (но 8 отмерло), в перми новых классов не появилось (но 4 отмерли). К триасу 3 класса отмерли, но 1 появился (и число классов достигло фанерозойского минимума, если не считать кембрия), в юре и мелу был небольшой прирост. В кайнозое изменений не произошло. Аналогичны данные и по классам растений; здесь также разделяются нижний палеозой, верхний палеозой, мезозой и кайнозой, лишь отмирания классов почти не происходило [7. С. 197].
Жизнь на Земле зародилась, когда на дне мелких теплых морей катархея, богатых сложными органическими веществами до аминокислот включительно, начали образовываться двойные сахаро-фосфатные спиральные нити высокополимерных нуклеиновых кислот с закрепленными на них последовательностями оснований (служащими «кодами» для синтеза белков), способные при некоторых условиях разворачиваться в одинарные спирали и синтезировать на каждой из них недостающую вторую спираль, т.е. порождать пару себе подобных (передавать им информацию о процедурах синтеза белков, закодированных последовательностями оснований). Такие полимеры, которые синтезировали белки, обеспечивающие им достаточно длительное самосохранение, по-видимому, уже можно считать первичными организмами.
Эти первичные микроорганизмы, возможно, питались имевшейся органикой небиологического происхождения, осуществляя, например, бескислородное разложение белков и аминокислот - гниение или углеродов - брожение (анологично питаются современные сапрофиты, поглощая через свои стенки клеток органику биологического происхождения: например, дрожжи при отсутствии кислорода сбраживают глюкозу до спирта и углекислоты). Развитие этих организмов, вероятно, исключило условия для дальнейшего самозарождения жизни, и с тех пор все живое появляется уже только от живого. В результате эволюции микроорганизмов (вынуждаемой нехваткой подходившей для пищи органики) у них появилась способность синтезировать необходимые для самосохранения органические молекулы из неорганических. Наиболее эффективным способом оказался фотосинтез - продуцирование органического вещества из углекислоты и воды под действием солнечного света (энергия света, поглощаемого пигментами, прежде всего зеленым хлорофиллом, расходуется на расщепление молекул воды, кислород выделяется в атмосферу, а водород вместе с углекислотой идет на образование первичного органического продукта - фосфоглицериновой кислоты).
Первыми фотосинтезирующими растениями были, по-видимому, микроскопические синезеленые водоросли цианофиты, у которых хлорофилл рассеян в виде мелких зерен по плазме клеток (у более развитых растений он сосредоточен в специальных тельцах - хлоропластах), и имеется еще синий пигмент фикоцианин. Эти водоросли похожи на бактерий тем, что в их клетках трудно различить ядра, и размножаются они только делением.
Наиболее древние остатки жизнедеятельности организмов, найденные в Трансваале в породах серии Свазиленд возрастом 3.1-3.4 млрд. лет, были тщательно изучены Э. Баргхорном и Дж. Шопфом. Они представляют собой микроскопические изолированные палочки длиной 0.45-0.7 мк. и диаметром 0.18-0.32 мк., имеющие двухслойные оболочки толщиной 0.045 мк.; там же обнаружены нитеподобные образования, а также микроскопические шаровидные, дисковидные и многоугольные оболочки одноклеточных водорослей акритархи. Почти столь же древними (более 2.9 млрд. лет) являются обызвествленные продукты жизнедеятельности цианофитов и бактерий - прикрепленные ко дну столбчатые строматолиты и неприкрепленные округлые онколиты, найденные в известковых прослоях зеленокаменных пород системы Булавайо в Южной Родезии и описанные А. Мак-Грегором еще в 1940 г. [7. С. 204]
Один из наиболее древних следов животных, найден А.М. Лейтесом в среднепротерозойских слоях Удоканской серии Забайкалья. Он представляет собой приуроченные к строматолитовым породам тонкостенные трубочки длиной до 2.5 см, диаметром до 4 мм, имеющие четырехгранное сечение с округленными углами и оставшиеся, по-видимому, от червей. По-видимому, к началу рифея кислорода было уже достаточно для развития примитивных животных. В породах среднего и верхнего рифея найдены следы роющих животных и трубочки червей. В породах венда обнаружены уже многочисленные отпечатки разнообразных бесскелетных животных, а начало кембрия маркируется массовым появлением скелетообразующих организмов.
Палеонтологические данные позволили воссоздать грандиозную и стройную картину эволюции форм жизни в фанерозое, не оставляющую никаких сомнений в естественности этого процесса. Проследим фанерозойскую эволюцию ряда важнейших типов организмов.
Начнем с растений, которые осуществляют фотосинтез первичного органического вещества и потому являются первоосновой всей жизни. Водоросли появились еще в начале архея (или, может быть, даже в катархее) и с тех пор благополучно существуют до настоящего времени преимущественно в виде одноклеточных форм. Доля водорослей в общей массе живых организмов на Земле в наше время составляет меньше одной тысячной (около 1.7 млрд. т из общей суммы в 2-3 триллиона т), но зато они чрезвычайно быстро размножаются - их клетки делятся каждые 1-2 суток, и в результате их продукция, около 550 млрд. т в год, составляет основную долю продукции биомассы на Земле в целом.
Первые наземные растения появились, вероятно, в ордовике или даже в кембрии, но достоверные их следы найдены лишь в силуре. Это были самые примитивные из высших растений типа сосудистых - псилофиты, мелкие растения без корней и листьев, а также первые плауны. В девоне появились представители двух других подтипов сосудистых растении - клинолистных (хвощей) и папоротникообразных (из последних - пока лишь наиболее примитивные растения класса папоротников а также отряда семенных папоротников птеридосперм из класса голосемянная), причем крупные древовидные формы плаунов хвощей и папоротников образовали первые леса нашей планеты. В карбоне обширные участки суши покрыли огромные заболоченные леса из гигантских плаунов лепидодендронов и сигиллярий, хвощей каламитов, древовидных папоротников и птеридосперм, а также новых отрядов вечнозеленых голосемянных кордаитов, а к концу периода - первых хвойных растении и гинкго. В перми появились бенеттиты, плауны пришли в упадок; на территориях Гондваны господствовала специфическая растительность во главе с голосемянными глоссоптерисами.
Триас и юра - это время расцвета голосемянных растений отрядов бенеттитов, хвойных, гинкго и цикадовых (саговников); птеридоспермы и кордаиты вымерли. В нижней половине мела, а возможно даже в юре, появились первые покрытосемянные (цветковые) растения, сначала двудольные. Во второй половине мела появились также однодольные цветковые растения, а двудольные образовали уже леса (кленовые и дубовые). Они вытеснили ряд отрядов голосемянных - бенеттиты вымерли, цикадовые и гинкго пришли в упадок. Наступивший затем кайнозой - эра расцвета цветковых растений.
Рассмотренная эволюция форм растений от водорослей до цветковых ттооисходила по линии уменьшения роли двуполых гаплоидных поколении, в телах которых (гаметофитах) клетки содержат одинарные наборы хромосом, и увеличений роли бесполых диплоидных поколений, в телах которых клетки содержат двойные наборы хромосом. Так, у многоклеточной водоросли улотрикс имеется только 1 диплоидная клетка, остальные - гаплоидны. У покрытосемянных, наоборот, все растение гаплоидно а гаметофит состоит лишь из нескольких клеток в тканях цветка; эволюционная выгода этого заключается как в том, что для размножения цветковых не требуется внешней влаги, так и в меньшей чувствительности диплоидных организмов к влиянию рецессивных генов (таких, которые действуют лишь при наличии их в обоих наборах хромосом в диплоидной клетке). В наше время наземные растения, преимущественно цветковые и хвойные, составляют 98-99% от 2-3 триллионов т биомассы на Земле, но растут они медленно - их годичная продукция составляет всего около 50 млрд. т (вдесятеро меньше продукции водорослей в океане), а средний возраст (очевидно, возраст деревьев в лесах) - 40-60 лет [7. С. 211].
Переходя к эволюции животных, отметим, что сведения палеонтологии о времени появления, расцвета и вымирания тех или иных групп животных вместе с данными о степени сложности их устройства, способах размножения и особенностях развития их зародышей позволяют судить о происхождении этих групп. Современные представления о происхождении основных типов животных (базирующиеся на принципе монофилии, т.е. на предположении о наличии единого корня происхождения каждой группы организмов) показаны на рис. 1. Согласно этим представлениям, одноклеточные простейшие животные породили как тупиковую ветвь губок, так и прогрессивную ветвь кишечнополостных, от которой пошли два главных ствола - первичноротых и вторичноротых. На первом из этих стволов промежуточным этапом были низшие черви, от которых пошли ветви немертин, плеченогих, мшанок, форонид, моллюсков и через кольчатых червей, наконец, членистоногие. От другого ствола поочередно отделялись ветви щетинкочелюстных червей, иглокожих, погонофор, кишечнодышащих, оболочников, головохордовых и, наконец, позвоночных.
Ряд типов животных - по крайней мере простейшие, кишечнополостные и некоторые низшие черви - появился, хотя бы в небольших популяциях, еще в среднем протерозое. Число семейств - одна из возможных количественных характеристик разнообразия форм данного типа организмов, свидетельствующая о темпах его эволюции: в периоды быстрой эволюции разнообразие форм должно возрастать.
К сожалению, мы не имеем возможности остановиться на истории развития каждого из типов животных - напомним, что всего имеется 23 типа!
Поэтому о первичноротых мы скажем лишь то, что все их типы появились, по-видимому, еще в докембрии. Нам придется оставить в стороне такие интересные и важные вопросы, как, например, связанная с эволюцией кишечно-полостных история коралловых рифов или же охватывающая весь фанерозой история «приматов моря» - головоногих моллюсков (не раз порождавших и массовые формы, и таких гигантов, как наутилусы ордовика с 6-метровыми прямыми раковинами, меловые аммониты со спиральными раковинами 3-метрового диаметра и современные гигантские «кракены» - глубоководные кальмары архитевтисы длиной до 15-20 м.).
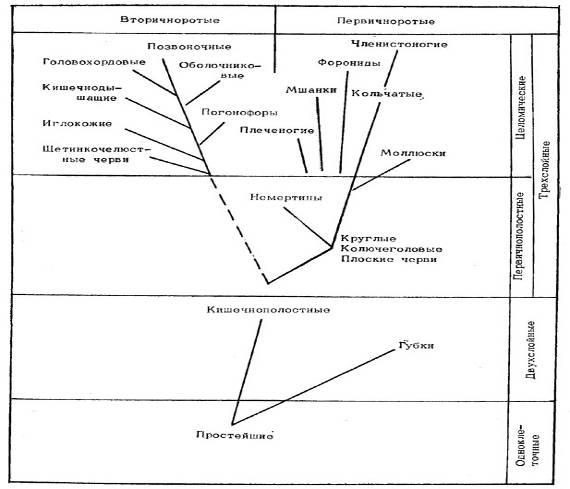
Рис. 1. Происхождение основных типов животных.
Из вторичноротых рассмотрим лишь историю позвоночных. Первые их представители появились в кембрии, это так называемые панцирные рыбы - небольшие рыбообразные донные животные без челюстей и плавников, покрытые панцирем из костных пластинок на голове и толстых чешуи на туловище; они еще не были настоящими рыбами - из современных позвоночных животных к ним ближе всего бесчерепные (ланцетники) и бесчелюстные - круглоротые (миноги и миксины). В силуре появились, а в девоне получили широкое развитие уже и настоящие рыбы, как хрящевые (сначала панцирные, а затем и беспанцирные акулы, благоденствующие с тех пор по настоящее время), так и костные, уже в середине девона поделившиеся на лопастеперых (двоякодышащих и кистеперых) и лучеперых, сначала ганоидных в триасе, а с юры и костистых, имеющих абсолютное господство среди рыб в настоящее время. Девон был временем первых попыток рыб выбраться на сушу; двоякодышащие рыбы в этом не преуспели, а от кистеперых пошли земноводные животные (амфибии), которые осваивали сушу одновременно с насекомыми.
Первые настоящие наземные позвоночные - это пресмыкающиеся (рептилии), у которых размножение и развитие происходят на суше. История рептилий перми и мезозоя, этих двухсот миллионов лет господства чудовищ на нашей планете, является, вероятно, самой драматической главой истории жизни на Земле до появления человека. Современное представление о пресмыкающихся, - как о холоднокровных животных с чешуйчатой кожей, плохо приспособленных к среде с низкой или резко колеблющейся температурой, возможно, не следует распространять на динозавров. Для выяснения этого вопроса Роберт Беккер (1975 г.) использовал три признака, отличающих теплокровных животных от холоднокровных:
1) наличие в костях большого количества кровеносных сосудов, а также каналов, в которых происходит быстрый обмен фосфатом кальция, необходимый для нормального функционирования мышц и нервов;
2) малое отношение биомасс хищников и их жертв (порядка 1-5% против десятков процентов у холоднокровных животных);
3) способность существовать в условиях холодного климата [8. С. 92].
По всем трем признакам установлено, что если раннепермские хищные пеликозавры (например, диметродоны) еще были холоднокровными, то уже позднепермские терапсиды и позднетриасовые текодонты были теплокровными (отношения хищник-жертва порядка 10-15%); у динозавров признаки теплокровности выражены даже ярче, чем у современных млекопитающих (отношение хищник-жертва всего 1-3%, так что находки костей хищных динозавров чрезвычайно редки). Динозавры, особенно жившие в высоких широтах, вероятно, имели и волосяные покровы.
Таким образом, эволюция форм организмов происходит, во-первых, вследствие мутаций (вызываемых физико-химическими воздействиями случайных изменений в нуклеинокислотных носителях наследственности) и, во-вторых, вследствие отбора тех мутантов, которые оказываются способными к адаптации (т.е. к приспособлению) к окружающей их живой и неживой природе. В большинстве случаев способными к адаптации оказываются мутанты с полезными усложнениями организмов; полезные упрощения возникают гораздо реже, и жизнь в целом эволюционирует от простого к сложному. Иногда же возникают особенно удачные формы организмов, столь хорошо приспособленные не только к среде, окружавшей их в период образования, но и к широкому спектру ее изменений, что им удается просуществовать чрезвычайно долго, до сотен миллионов лет включительно. Иначе говоря, высокая мера адаптированности приводит к низкой мере эволюционирования (и наоборот) [8. С. 89].
Человеческое общество оказывает все усиливающееся влияние на развитие жизни на Земле, которое началось в форме охоты, рыболовства и истребления опасных для человека животных и вытеснения дикой жизни из населенных районов, а в наши дни идет по пути создания и разведения культурных сортов растений и пород животных и может, в конце концов, привести почти к стопроцентному управлению развитием всех форм жизни.
Это может касаться и вида Homo sapiens, развитие которого определяется не столько биологическими, сколько социальными факторами - прежде всего уровнем производительных сил общества и производственными отношениями его членов (и такими надстройками, как складывающаяся стихийно или формируемая сознательно общественная мораль, которая, например, в прошлые столетия вследствие высокой детской смертности поощряла многодетность; в последние же десятилетия при снижении детской смертности, но создаваемых урбанизацией жилищных ограничениях это отношение изменяется).
Заключение
В современной теории эволюции значительное место занимают вопросы происхождения жизни на Земле, ибо в рамках теории эволюции происхождение жизни рассматривают в качестве изначального процесса образования живых форм (необиогенеза) из неорганических веществ, т.е. в виде химической эволюции. В наше время абиогенезу противопоставлена концепция постепенного возникновения и развития живого материала в течение длительного периода времени. Эта концепция получила название архебиоза или археогенеза. В соответствии с теорией Опарина жизнь является результатом исторического односторонне направленного развития в виде постепенного усложнения органических субъединиц и развития их в сложные системы, обладающие свойствами живого. Несмотря на остающуюся неопределенность в вопросе о происхождении жизни на Земле, не вызывает сомнения факт в естественности ее происхождения как определенного закономерного этапа развития материи.
1. Возникновение жизни на Земле подготовлено ходом эволюции неживой материи во Вселенной.
2. Существование жизни на Земле определяется постоянством действия космических факторов: мощностью и спектральным составом солнечного излучения, неизменностью основных характеристик орбиты Земли и ее осевого вращения, наличием магнитного поля и атмосферы планеты.
3. Развитие жизни на Земле во многом обусловлено плавными незначительными изменениями космических факторов; сильные изменения ведут к катастрофическим последствиям: космические лучи и их рассмотрение как мутагенных факторов.
4. На определенном этапе своего развития жизнь становится фактором космического масштаба, оказывающим влияние на физико-химические характеристики основных оболочек планеты (например, состав и температуру атмосферы, гидросферы и верхних слоев литосферы).
Список используемой литературы
1. Барг О.А. Живое в едином мировом процессе. Изд-во Пермского университета 1993. – 227 с.
2. Вселенная, астрономия, философия. М.: изд. МГУ 1988. – 192 с.
3. Данилова В.С., Кожевников Н.И. Основные концепции современного естествознания: учебник, М.: Аспект-пресс, 2000 – 256 с.
4. Карпенков С.Х. Современное естествознание: учеб., М.: Академический проект 2003. – 560 с.
5. Найденыш В.М. Концепции современного естествознания: Учеб. пособие д/вузов, М.: Гардарики 2002, - 476 с.
6. Соколов Е.Ф. Концепции современного естествознания: Учеб. пособие д/вузов, М.: ВЛАДОС, 1999. – 232 с.
7. .Шкловский И.С. Вселенная, жизнь, разум – М.: Наука, 1997 г. – 320 с.
8. Щербаков В.П. Эволюция как сопротивление энтропии – М.: ЭКСМО, 2006 г. – 125 с.